

Esc2 orchestrates substrate-specific sumoylation by acting as a SUMO E2 cofactor in genome maintenance
- PMID: 33446573
- PMCID: PMC7849368
- DOI: 10.1101/gad.344739.120
Esc2 orchestrates substrate-specific sumoylation by acting as a SUMO E2 cofactor in genome maintenance
Abstract
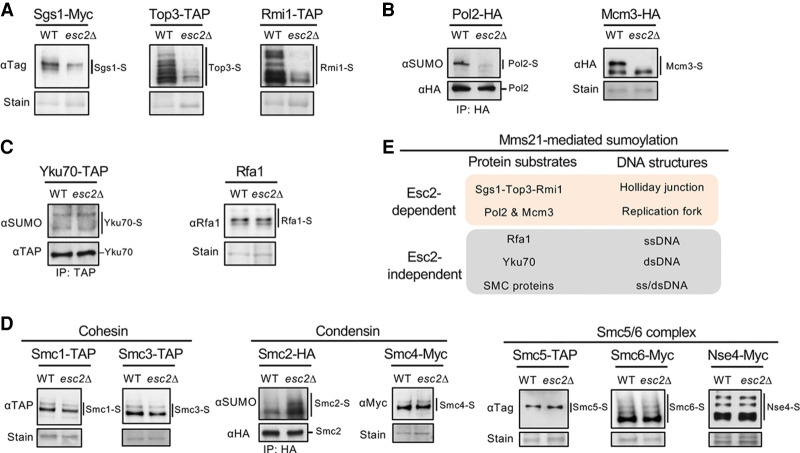
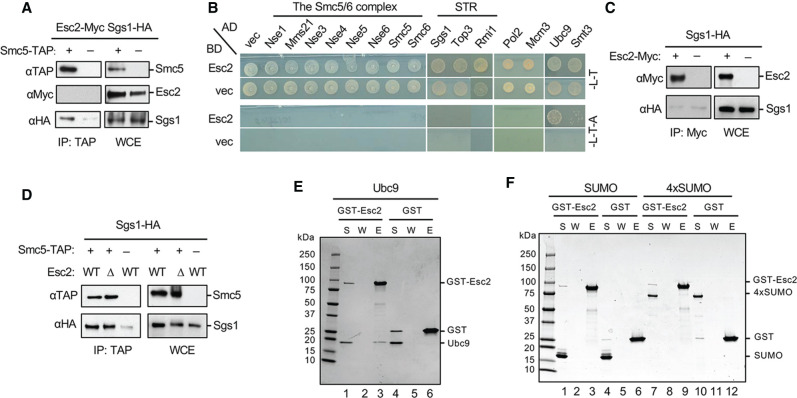
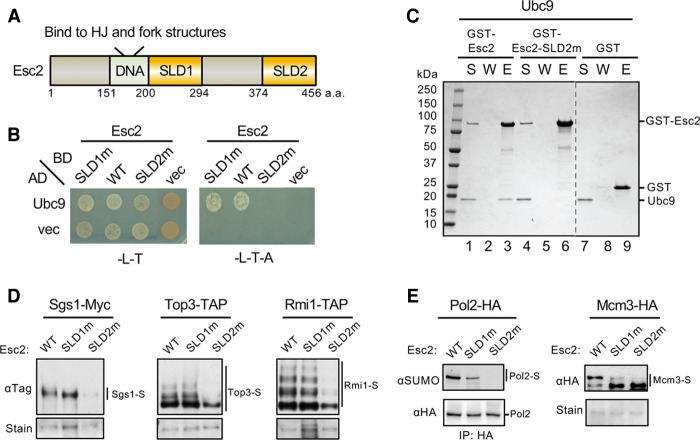
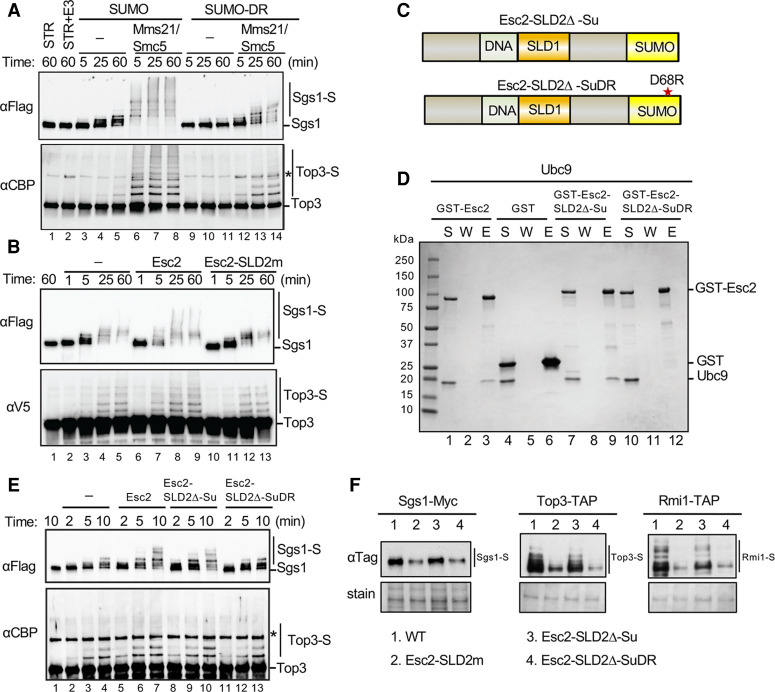
SUMO modification regulates diverse cellular processes by targeting hundreds of proteins. However, the limited number of sumoylation enzymes raises the question of how such a large number of substrates are efficiently modified. Specifically, how genome maintenance factors are dynamically sumoylated at DNA replication and repair sites to modulate their functions is poorly understood. Here, we demonstrate a role for the conserved yeast Esc2 protein in this process by acting as a SUMO E2 cofactor. Esc2 is required for genome stability and binds to Holliday junctions and replication fork structures. Our targeted screen found that Esc2 promotes the sumoylation of a Holliday junction dissolution complex and specific replisome proteins. Esc2 does not elicit these effects via stable interactions with substrates or their common SUMO E3. Rather, we show that a SUMO-like domain of Esc2 stimulates sumoylation by exploiting a noncovalent SUMO binding site on the E2 enzyme. This role of Esc2 in sumoylation is required for Holliday junction clearance and genome stability. Our findings thus suggest that Esc2 acts as a SUMO E2 cofactor at distinct DNA structures to promote the sumoylation of specific substrates and genome maintenance.
Keywords: Esc2; SUMO E2; genome maintenance; homologous recombination.
© 2021 Li et al.; Published by Cold Spring Harbor Laboratory Press.
Figures
References
-
- Bermúdez-López M, Villoria MT, Esteras M, Jarmuz A, Torres-Rosell J, Clemente-Blanco A, Aragon L. 2016. Sgs1's roles in DNA end resection, HJ dissolution, and crossover suppression require a two-step SUMO regulation dependent on Smc5/6. Genes Dev 30: 1339–1356. 10.1101/gad.278275.116 - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases