

Aneuploidy in human eggs: contributions of the meiotic spindle
- PMID: 33449109
- PMCID: PMC7925012
- DOI: 10.1042/BST20200043
Aneuploidy in human eggs: contributions of the meiotic spindle
Abstract
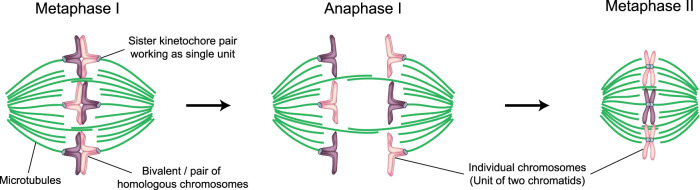
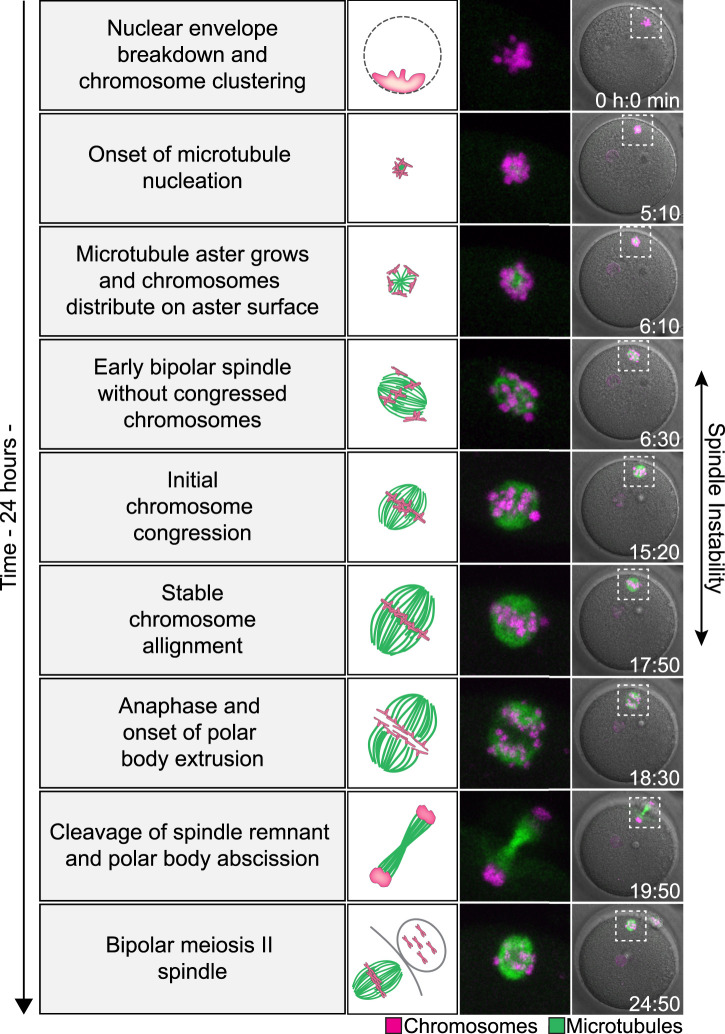
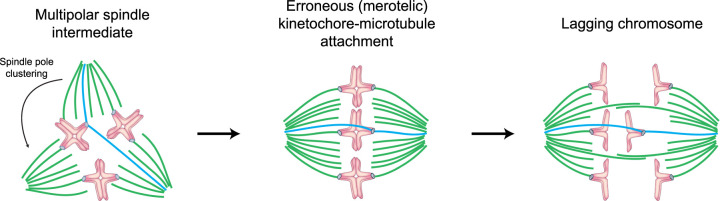
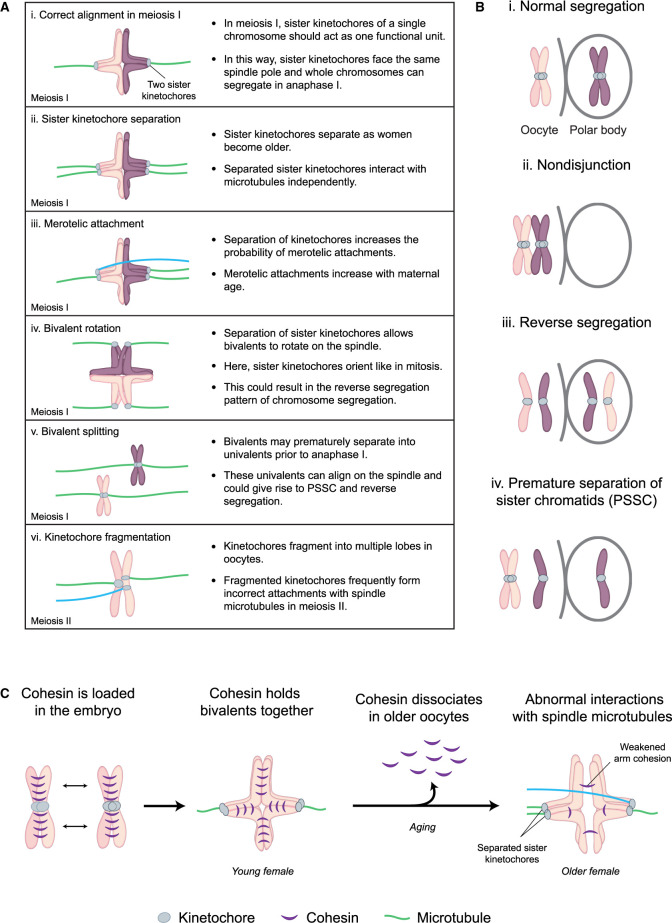
Human eggs frequently contain an incorrect number of chromosomes, a condition termed aneuploidy. Aneuploidy affects ∼10-25% of eggs in women in their early 30s, and more than 50% of eggs from women over 40. Most aneuploid eggs cannot develop to term upon fertilization, making aneuploidy in eggs a leading cause of miscarriages and infertility. The cellular origins of aneuploidy in human eggs are incompletely understood. Aneuploidy arises from chromosome segregation errors during the two meiotic divisions of the oocyte, the progenitor cell of the egg. Chromosome segregation is driven by a microtubule spindle, which captures and separates the paired chromosomes during meiosis I, and sister chromatids during meiosis II. Recent studies reveal that defects in the organization of the acentrosomal meiotic spindle contribute to human egg aneuploidy. The microtubules of the human oocyte spindle are very frequently incorrectly attached to meiotic kinetochores, the multi-protein complexes on chromosomes to which microtubules bind. Multiple features of human oocyte spindles favour incorrect attachments. These include spindle instability and many age-related changes in chromosome and kinetochore architecture. Here, we review how the unusual spindle assembly mechanism in human oocytes contributes to the remarkably high levels of aneuploidy in young human eggs, and how age-related changes in chromosome and kinetochore architecture cause aneuploidy levels to rise even higher as women approach their forties.
Keywords: aneuploidy; human oocyte; kinetochores; meiosis; spindle.
© 2021 The Author(s).
Conflict of interest statement
The authors declare that there are no competing interests associated with the manuscript.
Figures
References
-
- Centers for Disease Control and Prevention. Assisted Reproductive Technology Fertility Clinic Success Rates Report - 2017. 2017;17:105–16. https://www.cdc.gov/art/reports/2017/fertility-clinic.html.
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources