

Regulation of SIRT2 by Wnt/β-catenin signaling pathway in colorectal cancer cells
- PMID: 33450304
- PMCID: PMC7939112
- DOI: 10.1016/j.bbamcr.2021.118966
Regulation of SIRT2 by Wnt/β-catenin signaling pathway in colorectal cancer cells
Abstract
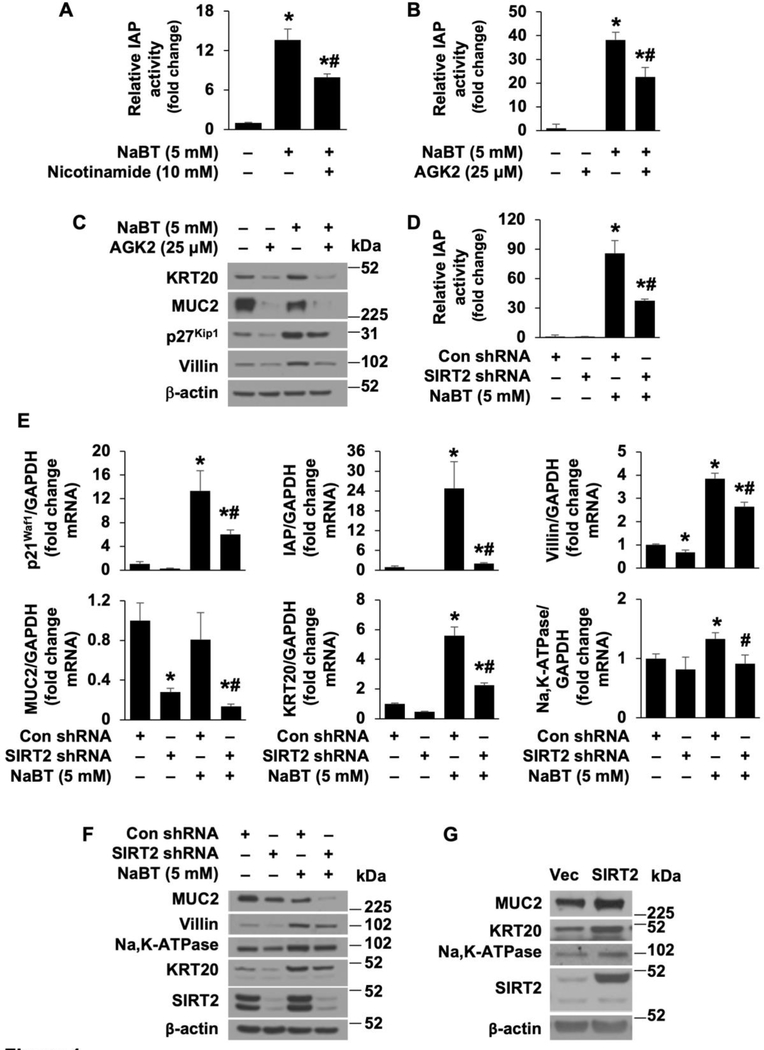
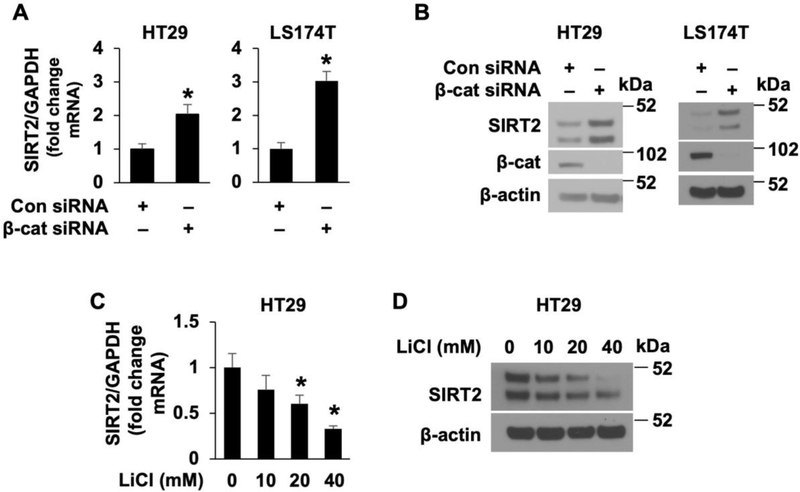
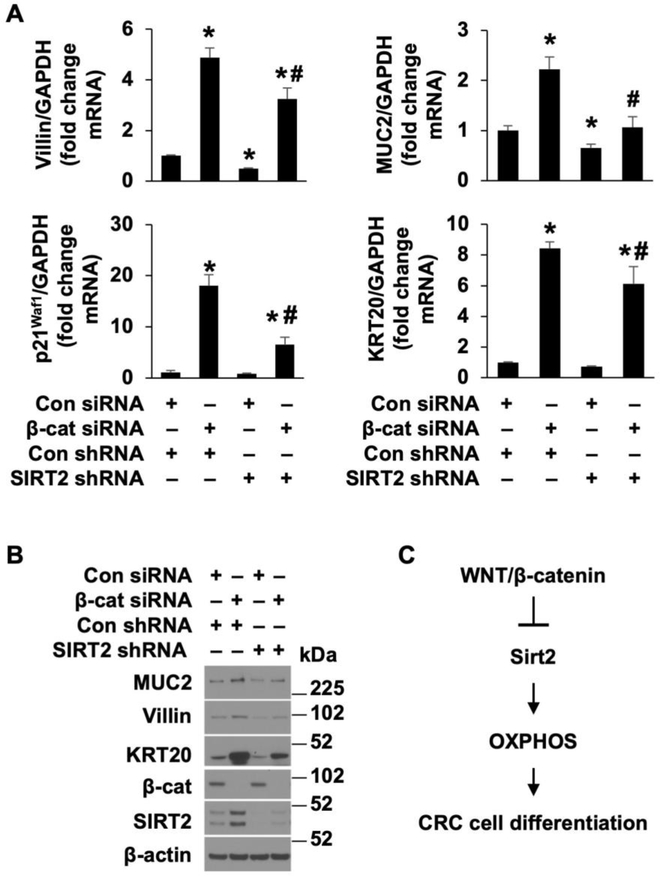
Activation of the Wnt/β-catenin pathway is one of the hallmarks of colorectal cancer (CRC). Sirtuin 2 (SIRT2) protein has been shown to inhibit CRC proliferation. Previously, we reported that SIRT2 plays an important role in the maintenance of normal intestinal cell homeostasis. Here, we show that SIRT2 is a direct target gene of Wnt/β-catenin signaling in CRC cells. Inhibition or knockdown of Wnt/β-catenin increased SIRT2 promoter activity and mRNA and protein expression, whereas activation of Wnt/β-catenin decreased SIRT2 promoter activity and expression. β-Catenin was recruited to the promoter of SIRT2 and transcriptionally regulated SIRT2 expression. Wnt/β-catenin inhibition increased mitochondrial oxidative phosphorylation (OXPHOS) and CRC cell differentiation. Moreover, inhibition of OXPHOS attenuated the differentiation of CRC cells induced by Wnt/β-catenin inhibition. In contrast, inhibition or knockdown of SIRT2 decreased, while overexpression of SIRT2 increased, OXPHOS activity and differentiation in CRC cells. Consistently, inhibition or knockdown or SIRT2 attenuated the differentiation induced by Wnt/β-catenin inhibition. These results demonstrate that SIRT2 is a novel target gene of the Wnt/β-catenin signaling and contributes to the differentiation of CRC cells.
Keywords: Cell differentiation; Cell signaling; Protein expression.
Copyright © 2021 Elsevier B.V. All rights reserved.
Conflict of interest statement
Declaration of interests
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
Figures




References
-
- Bahrami A, Amerizadeh F, ShahidSales S, Khazaei M, Ghayour-Mobarhan M, Sadeghnia HR, Maftouh M, Hassanian SM, Avan A, Therapeutic Potential of Targeting Wnt/beta-Catenin Pathway in Treatment of Colorectal Cancer: Rational and Progress, Journal of cellular biochemistry, (2017). - PubMed
-
- Gordon MD, Nusse R, Wnt signaling: multiple pathways, multiple receptors, and multiple transcription factors, J Biol Chem, 281 (2006) 22429–22433. - PubMed
-
- Nusse R, Clevers H, Wnt/beta-Catenin Signaling, Disease, and Emerging Therapeutic Modalities, Cell, 169 (2017) 985–999. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Miscellaneous
