

Systematic analysis of differential rhythmic liver gene expression mediated by the circadian clock and feeding rhythms
- PMID: 33452134
- PMCID: PMC7826335
- DOI: 10.1073/pnas.2015803118
Systematic analysis of differential rhythmic liver gene expression mediated by the circadian clock and feeding rhythms
Abstract
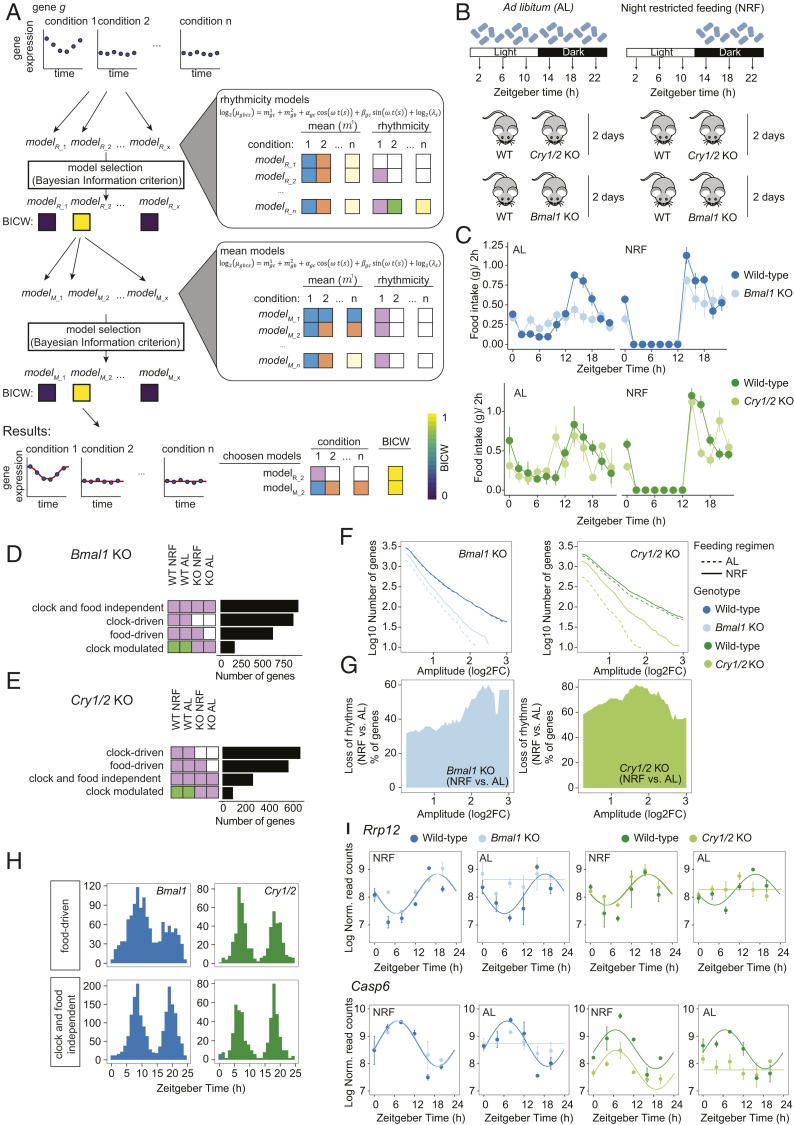
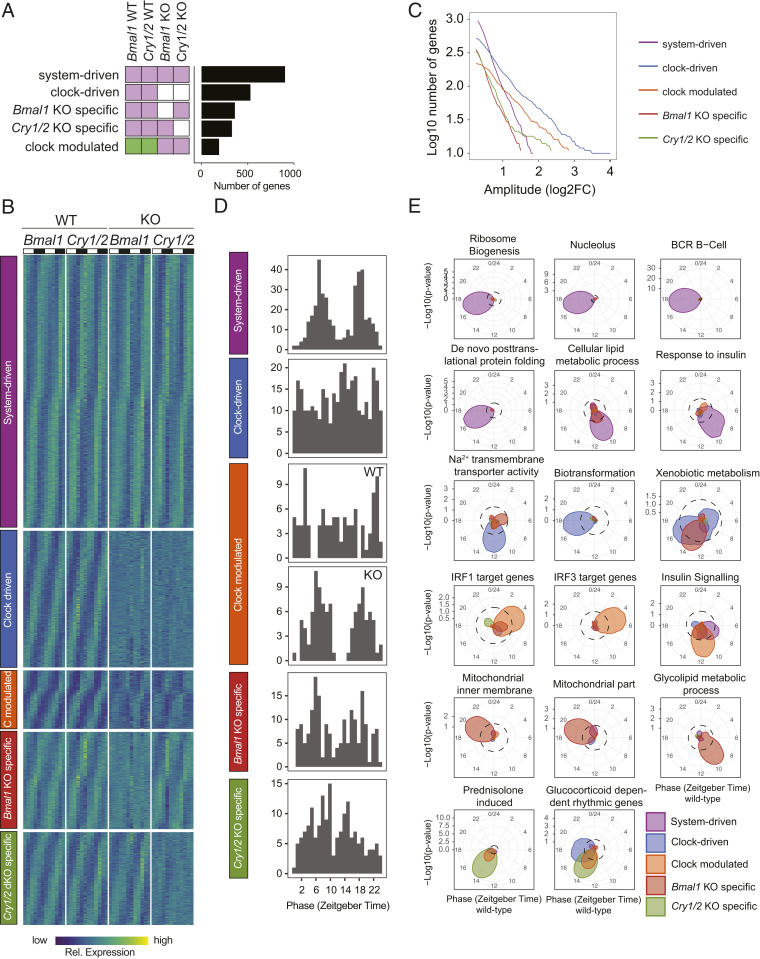
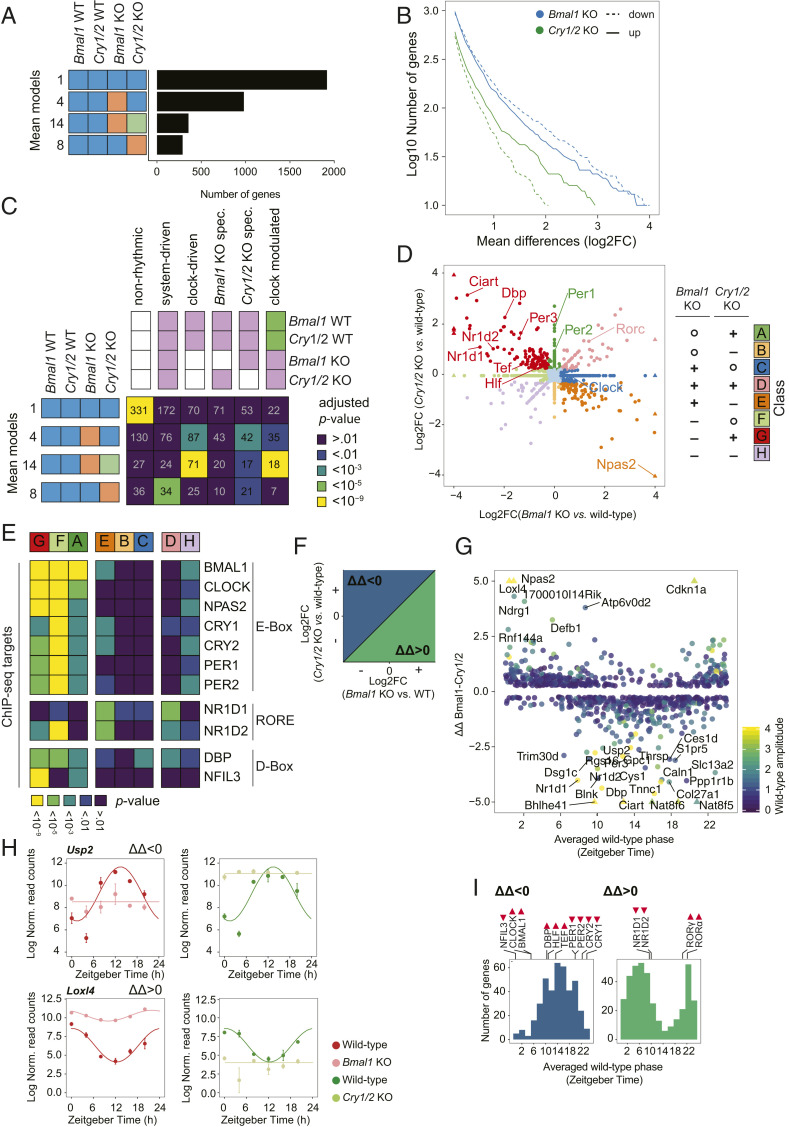
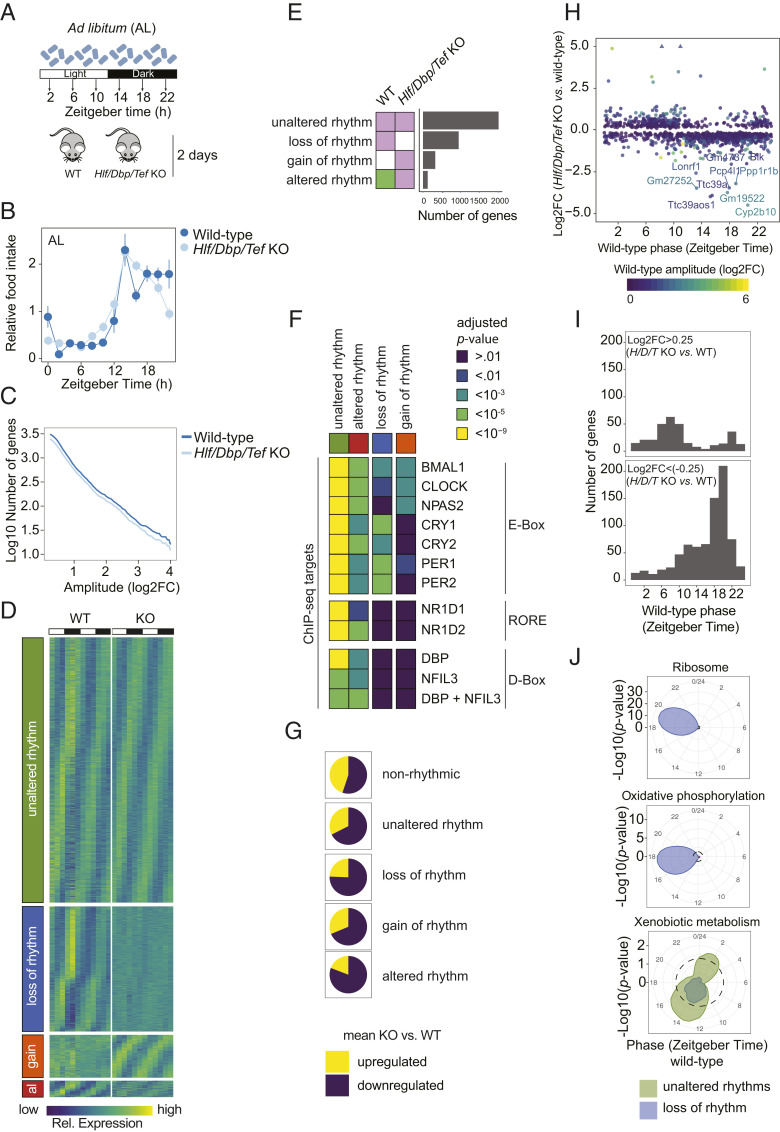
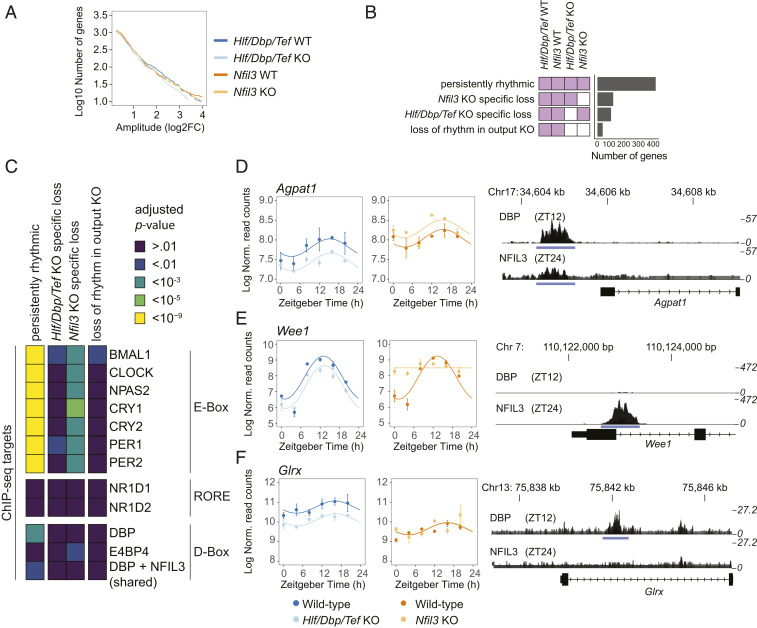
The circadian clock and feeding rhythms are both important regulators of rhythmic gene expression in the liver. To further dissect the respective contributions of feeding and the clock, we analyzed differential rhythmicity of liver tissue samples across several conditions. We developed a statistical method tailored to compare rhythmic liver messenger RNA (mRNA) expression in mouse knockout models of multiple clock genes, as well as PARbZip output transcription factors (Hlf/Dbp/Tef). Mice were exposed to ad libitum or night-restricted feeding under regular light-dark cycles. During ad libitum feeding, genetic ablation of the core clock attenuated rhythmic-feeding patterns, which could be restored by the night-restricted feeding regimen. High-amplitude mRNA expression rhythms in wild-type livers were driven by the circadian clock, but rhythmic feeding also contributed to rhythmic gene expression, albeit with significantly lower amplitudes. We observed that Bmal1 and Cry1/2 knockouts differed in their residual rhythmic gene expression. Differences in mean expression levels between wild types and knockouts correlated with rhythmic gene expression in wild type. Surprisingly, in PARbZip knockout mice, the mean expression levels of PARbZip targets were more strongly impacted than their rhythms, potentially due to the rhythmic activity of the D-box-repressor NFIL3. Genes that lost rhythmicity in PARbZip knockouts were identified to be indirect targets. Our findings provide insights into the diurnal transcriptome in mouse liver as we identified the differential contributions of several core clock regulators. In addition, we gained more insights on the specific effects of the feeding-fasting cycle.
Keywords: circadian clock; differential rhythmicity analysis; feeding–fasting cycle; liver metabolism; transcriptomics.
Copyright © 2021 the Author(s). Published by PNAS.
Conflict of interest statement
Competing interest statement: B.D.W., C.G., F.A., E.M, A.C., and F.G. were employees of Société des Produits Nestlé SA.
Figures
References
-
- Schibler U., et al. , Clock-talk: Interactions between central and peripheral circadian oscillators in mammals. Cold Spring Harb. Symp. Quant. Biol. 80, 223–232 (2015). - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
