

RADX controls RAD51 filament dynamics to regulate replication fork stability
- PMID: 33453169
- PMCID: PMC7935748
- DOI: 10.1016/j.molcel.2020.12.036
RADX controls RAD51 filament dynamics to regulate replication fork stability
Abstract
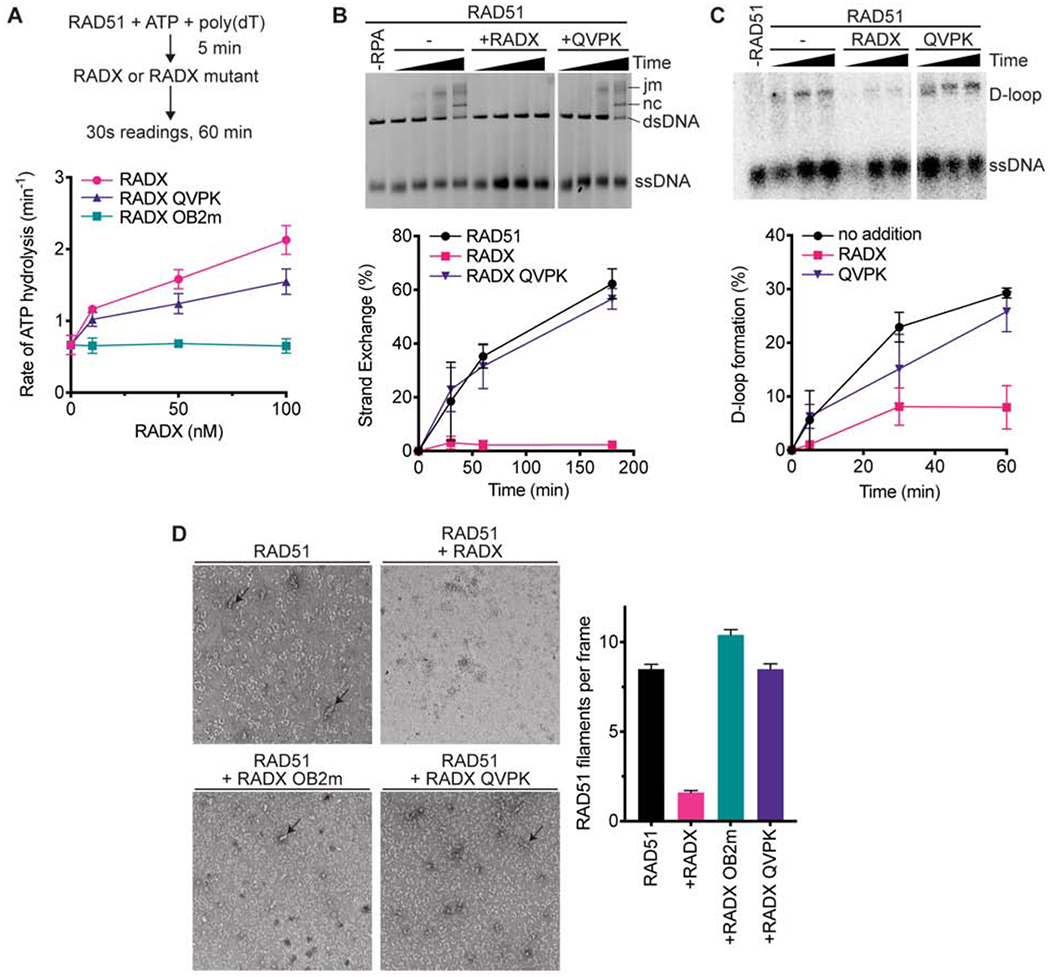
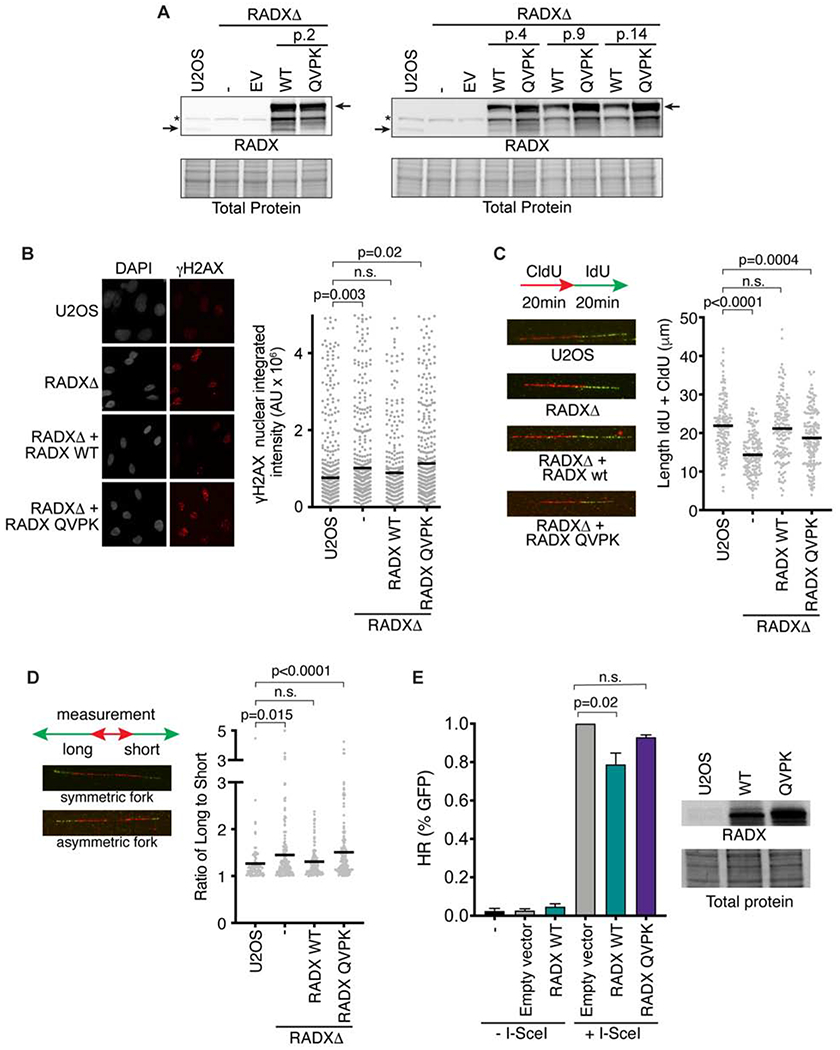
The RAD51 recombinase forms nucleoprotein filaments to promote double-strand break repair, replication fork reversal, and fork stabilization. The stability of these filaments is highly regulated, as both too little and too much RAD51 activity can cause genome instability. RADX is a single-strand DNA (ssDNA) binding protein that regulates DNA replication. Here, we define its mechanism of action. We find that RADX inhibits RAD51 strand exchange and D-loop formation activities. RADX directly and selectively interacts with ATP-bound RAD51, stimulates ATP hydrolysis, and destabilizes RAD51 nucleofilaments. The RADX interaction with RAD51, in addition to its ssDNA binding capability, is required to maintain replication fork elongation rates and fork stability. Furthermore, BRCA2 can overcome the RADX-dependent RAD51 inhibition. Thus, RADX functions in opposition to BRCA2 in regulating RAD51 nucleofilament stability to ensure the right level of RAD51 function during DNA replication.
Keywords: DNA curtain; DNA damage response; DNA repair; double-strand break; electron microscopy; fork reversal; replication stress.
Copyright © 2020 Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests The authors declare no competing interests.
Figures
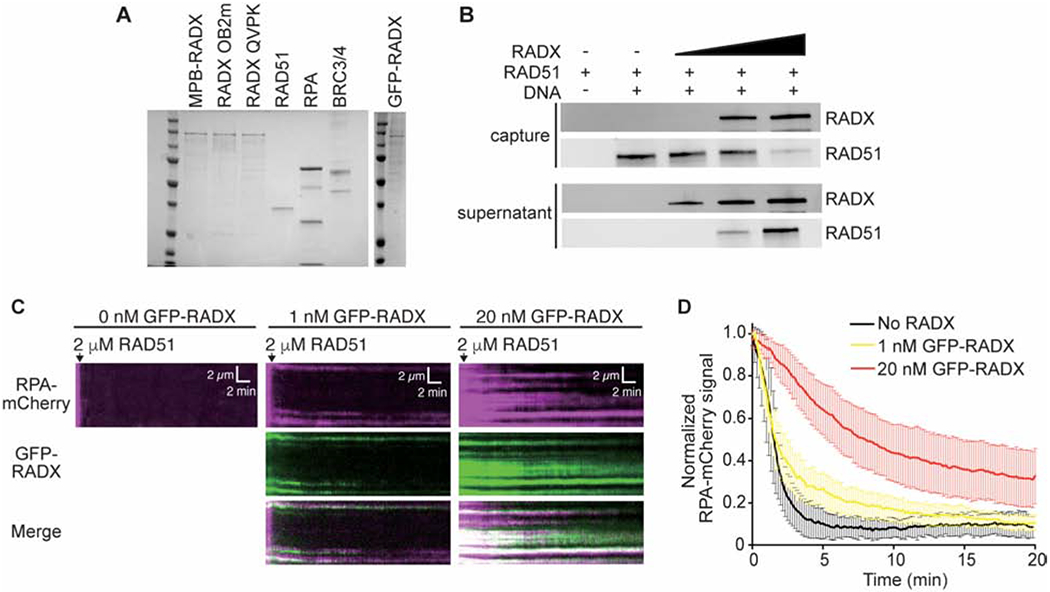
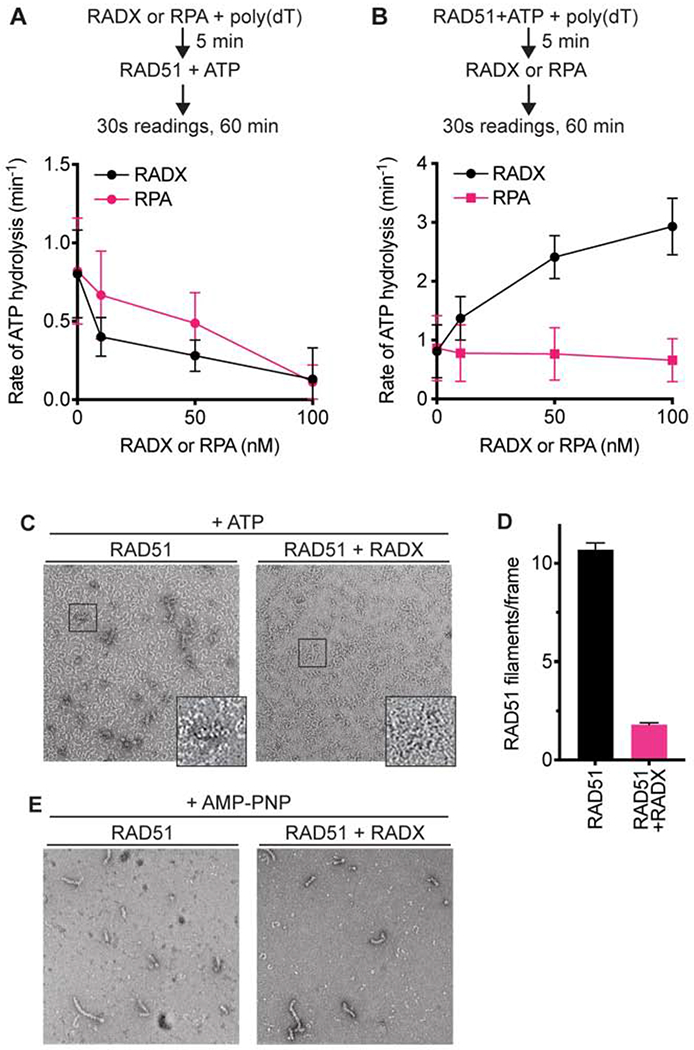
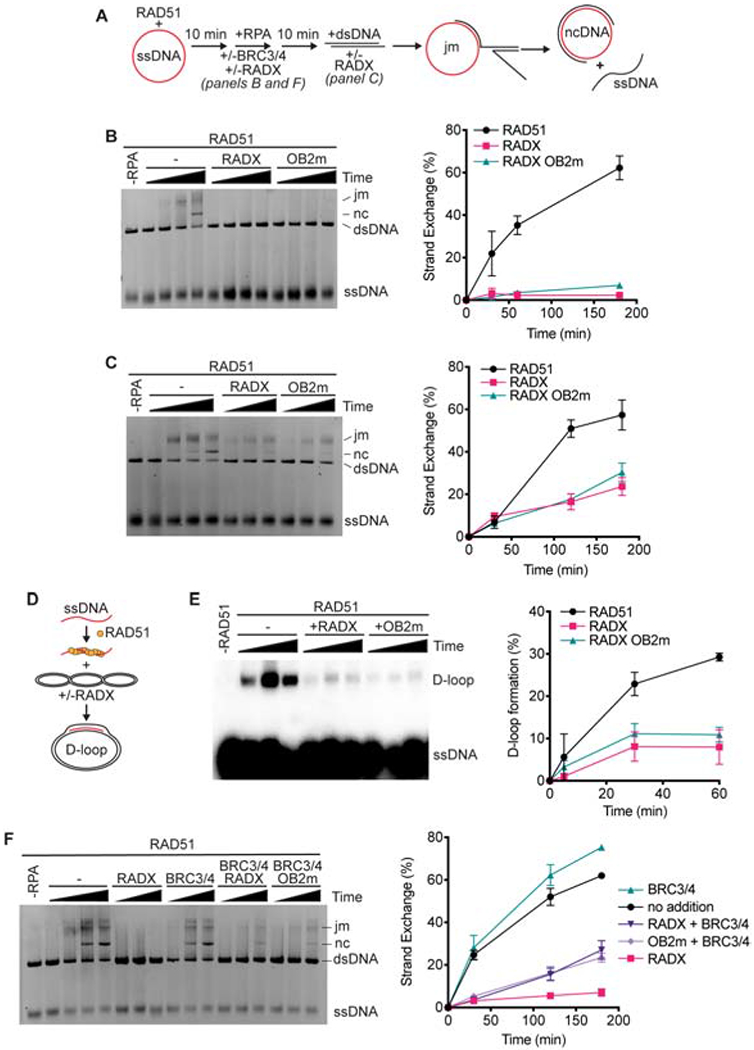
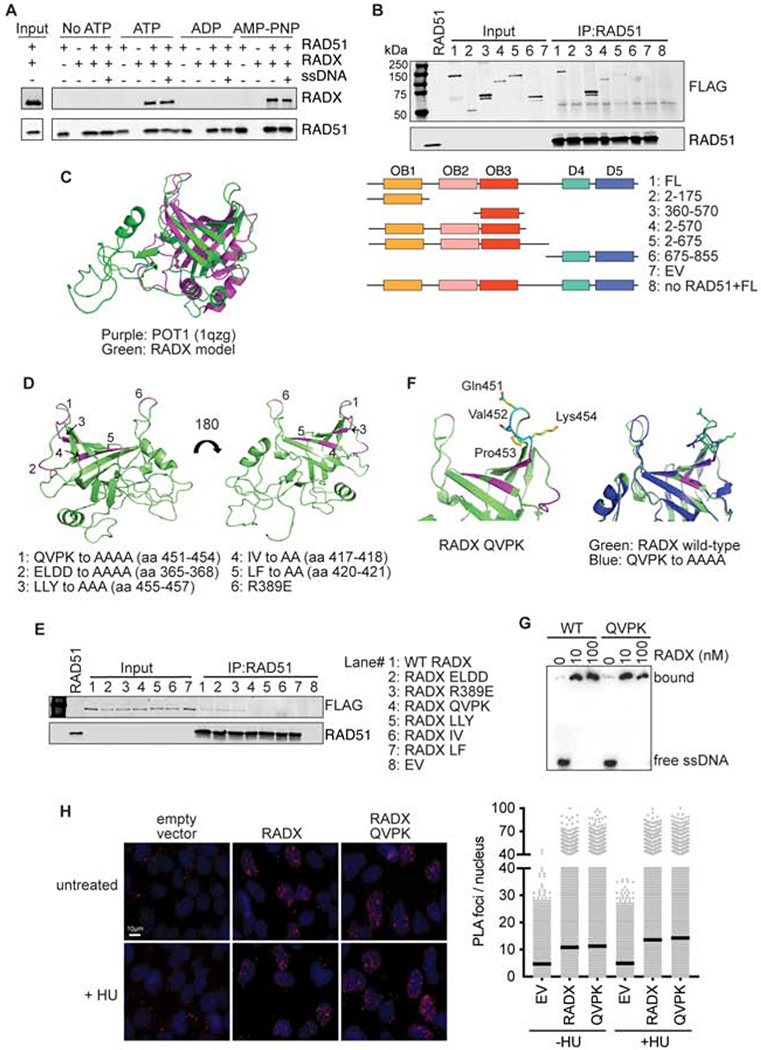
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
