

β-carotene oxygenase 2 deficiency-triggered mitochondrial oxidative stress promotes low-grade inflammation and metabolic dysfunction
- PMID: 33453359
- PMCID: PMC7946548
- DOI: 10.1016/j.freeradbiomed.2021.01.003
β-carotene oxygenase 2 deficiency-triggered mitochondrial oxidative stress promotes low-grade inflammation and metabolic dysfunction
Abstract
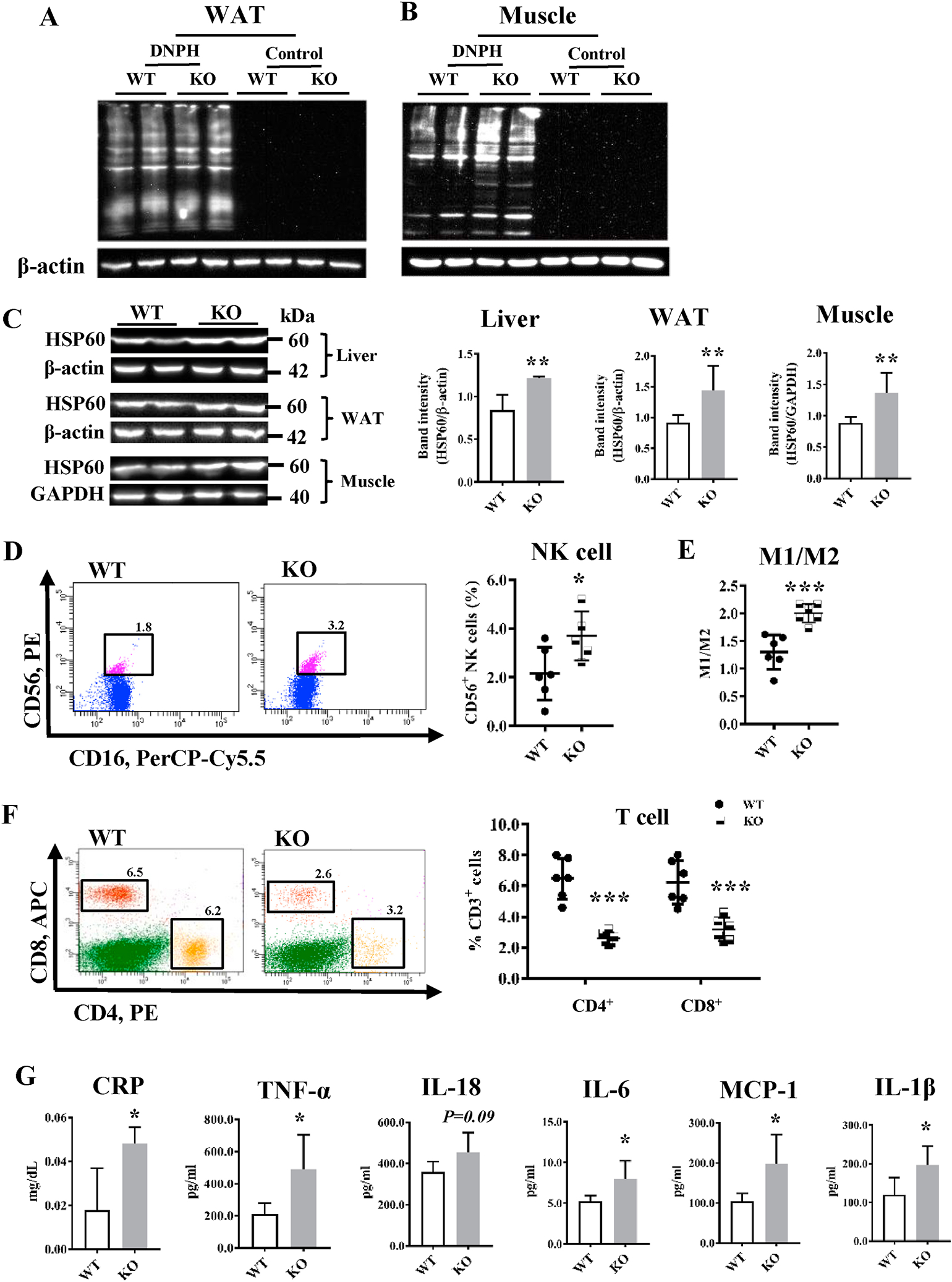
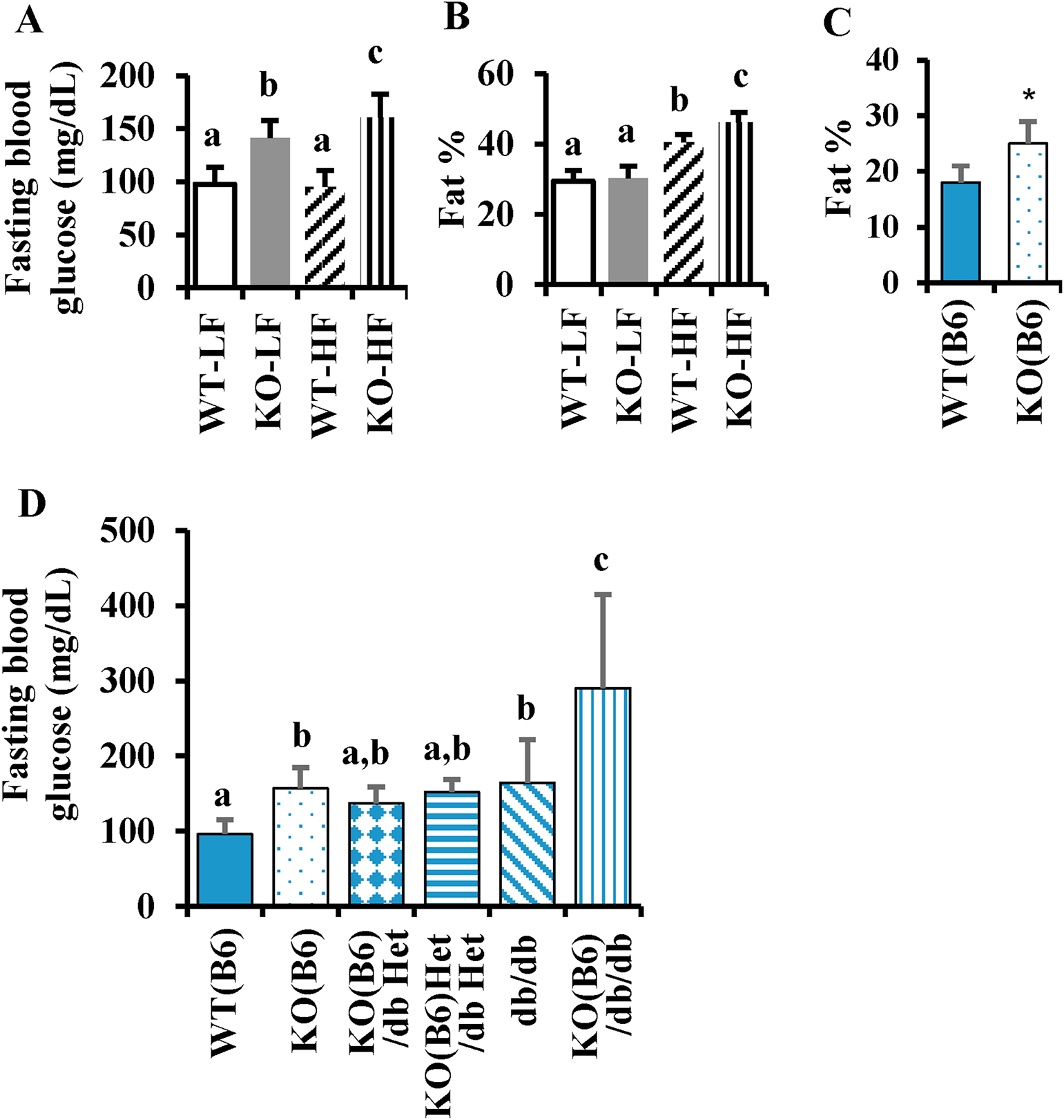
Low-grade inflammation is a critical pathological factor contributing to the development of metabolic disorders. β-carotene oxygenase 2 (BCO2) was initially identified as an enzyme catalyzing carotenoids in the inner mitochondrial membrane. Mutations in BCO2 are associated with inflammation and metabolic disorders in humans, yet the underlying mechanisms remain unknown. Here, we used loss-of-function approaches in mice and cell culture models to investigate the role of BCO2 in inflammation and metabolic dysfunction. We demonstrated decreases in BCO2 mRNA and protein levels and suppression of mitochondrial respiratory complex I proteins and mitochondrial superoxide dismutase levels in the liver of type 2 diabetic human subjects. Deficiency of BCO2 caused disruption of assembly of the mitochondrial respiratory supercomplexes, such as supercomplex III2+IV in mice, and overproduction of superoxide radicals in primary mouse embryonic fibroblasts. Further, deficiency of BCO2 increased protein carbonylation and populations of natural killer cells and M1 macrophages, and decreased populations of T cells, including CD4+ and/or CD8+ in the bone marrow and white adipose tissues. Elevation of plasma inflammatory cytokines and adipose tissue hypertrophy and inflammation were also characterized in BCO2 deficient mice. Moreover, BCO2 deficient mice were more susceptible to high-fat diet-induced obesity and hyperglycemia. Double knockout of BCO2 and leptin receptor genes caused a significantly greater elevation of the fasting blood glucose level in mice at 4 weeks of age, compared to the age- and sex-matched leptin receptor knockout. Finally, administration of Mito-TEMPO, a mitochondrial specific antioxidant attenuated systemic low-grade inflammation induced by BCO2 deficiency. Collectively, these findings suggest that BCO2 is essential for mitochondrial respiration and metabolic homeostasis in mammals. Loss or decreased expression of BCO2 leads to mitochondrial oxidative stress, low-grade inflammation, and the subsequent development of metabolic disorders.
Keywords: Diabetes; Human; Macrophage; Mitochondrial respiratory supercomplex assembly; Natural killer cell; Superoxide.
Copyright © 2021 Elsevier Inc. All rights reserved.
Conflict of interest statement
Competing interests
No competing interests declared
Figures





References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
