

Hydrogen/deuterium exchange memory NMR reveals structural epitopes involved in IgE cross-reactivity of allergenic lipid transfer proteins
- PMID: 33453986
- PMCID: PMC7762950
- DOI: 10.1074/jbc.RA120.014243
Hydrogen/deuterium exchange memory NMR reveals structural epitopes involved in IgE cross-reactivity of allergenic lipid transfer proteins
Abstract
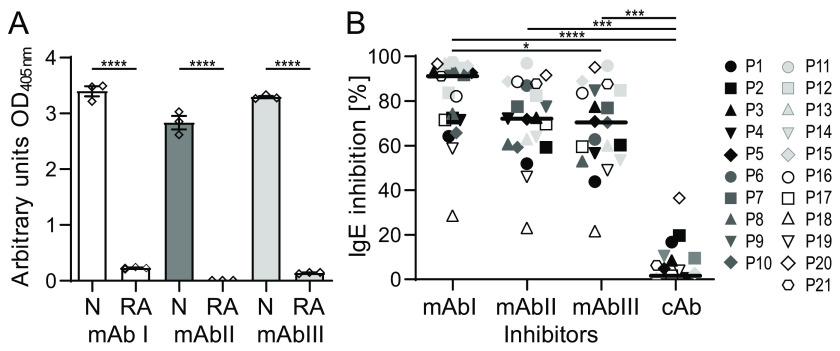
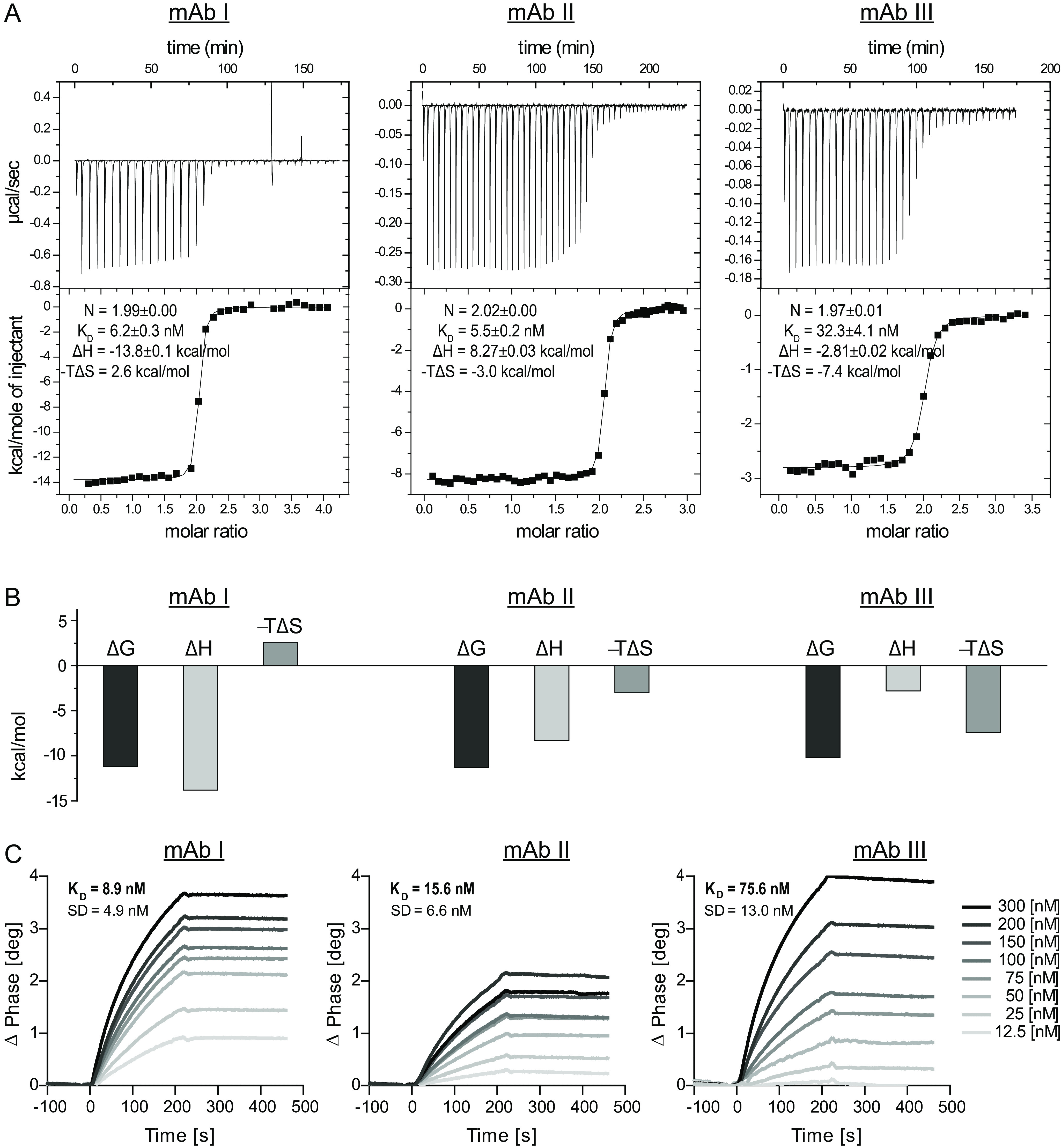
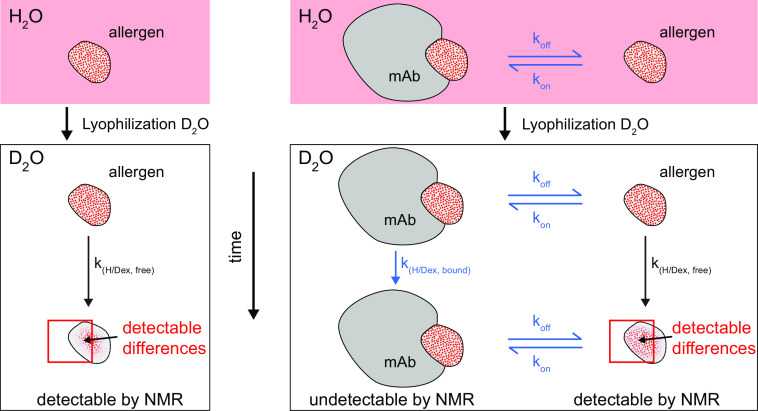
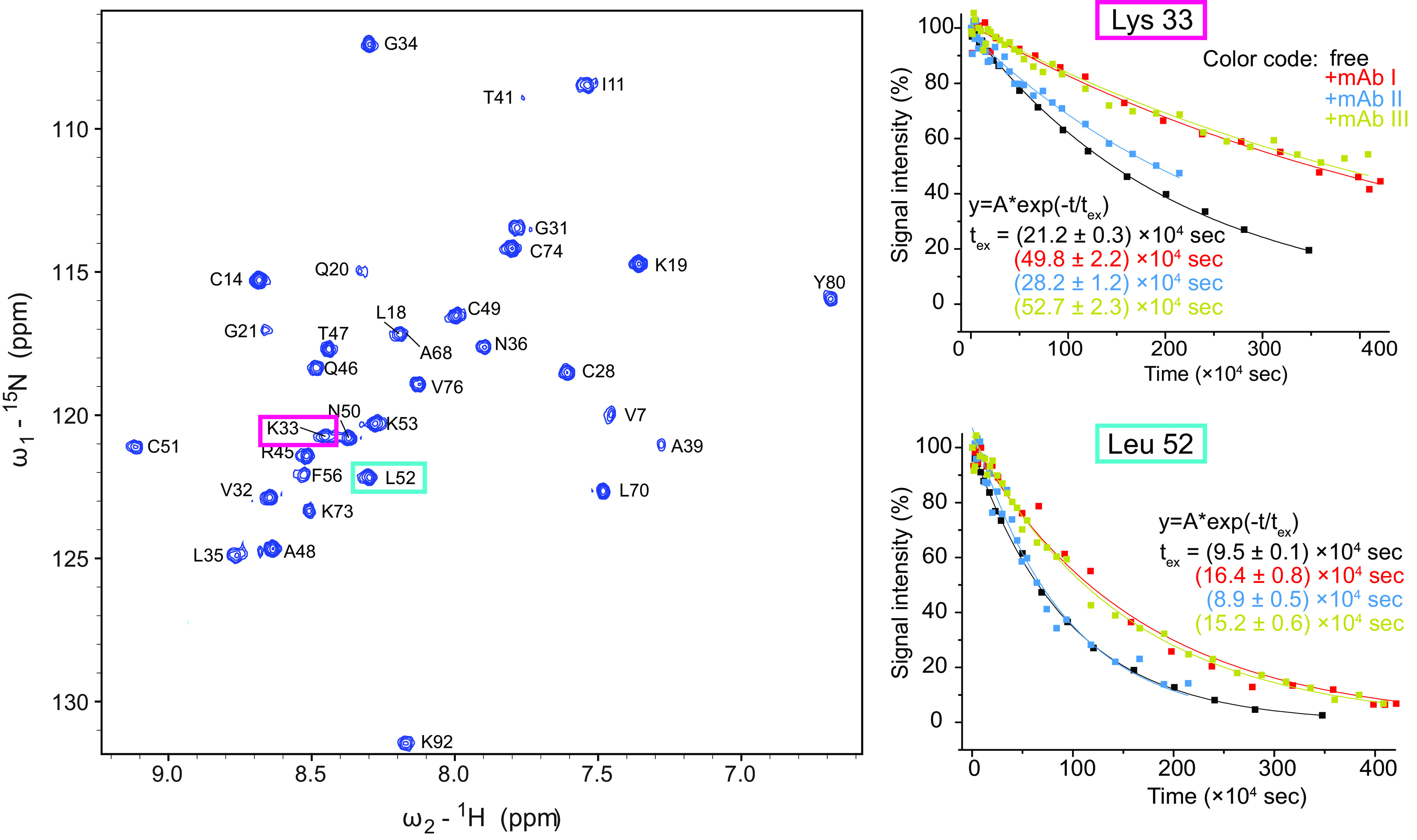
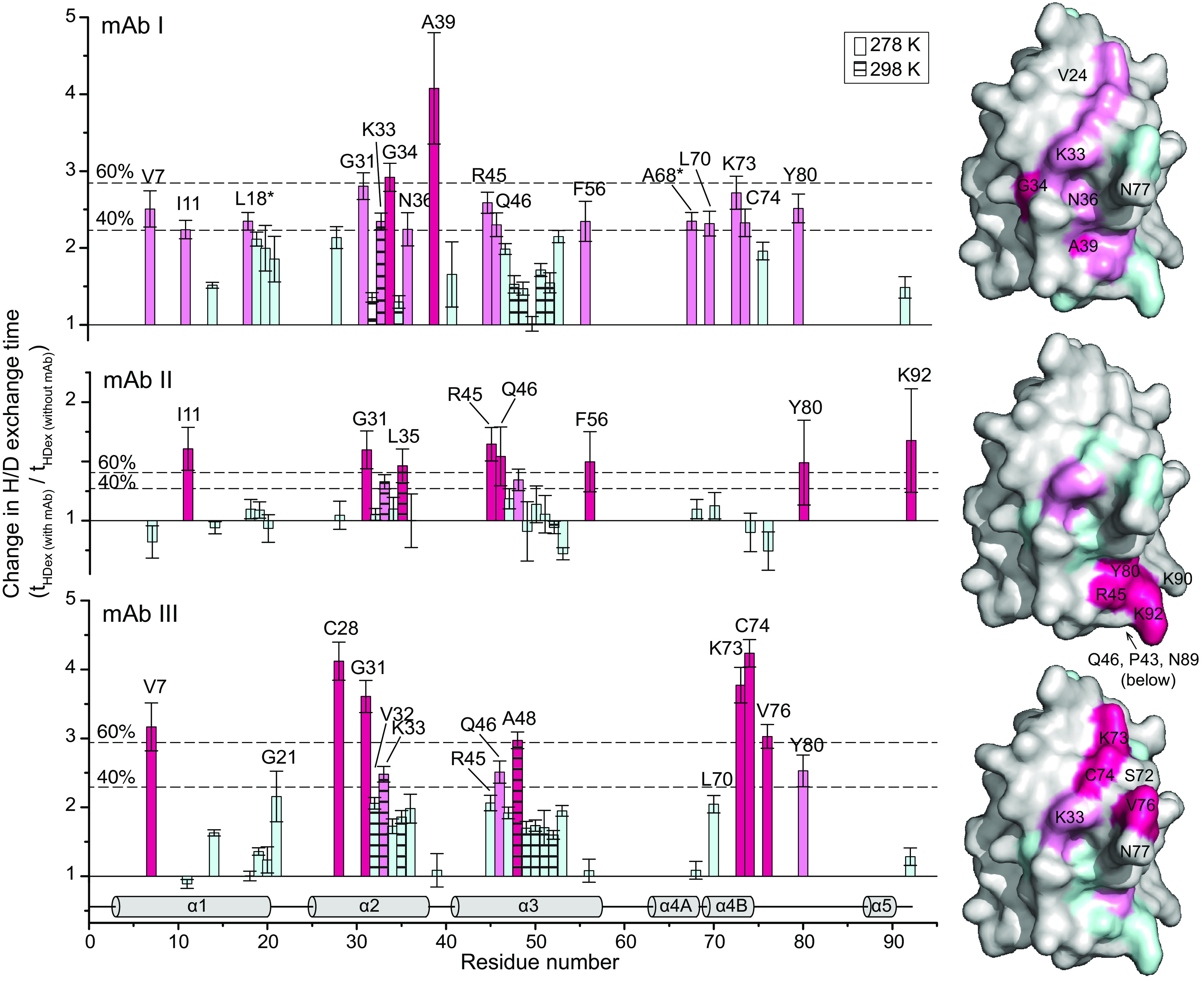
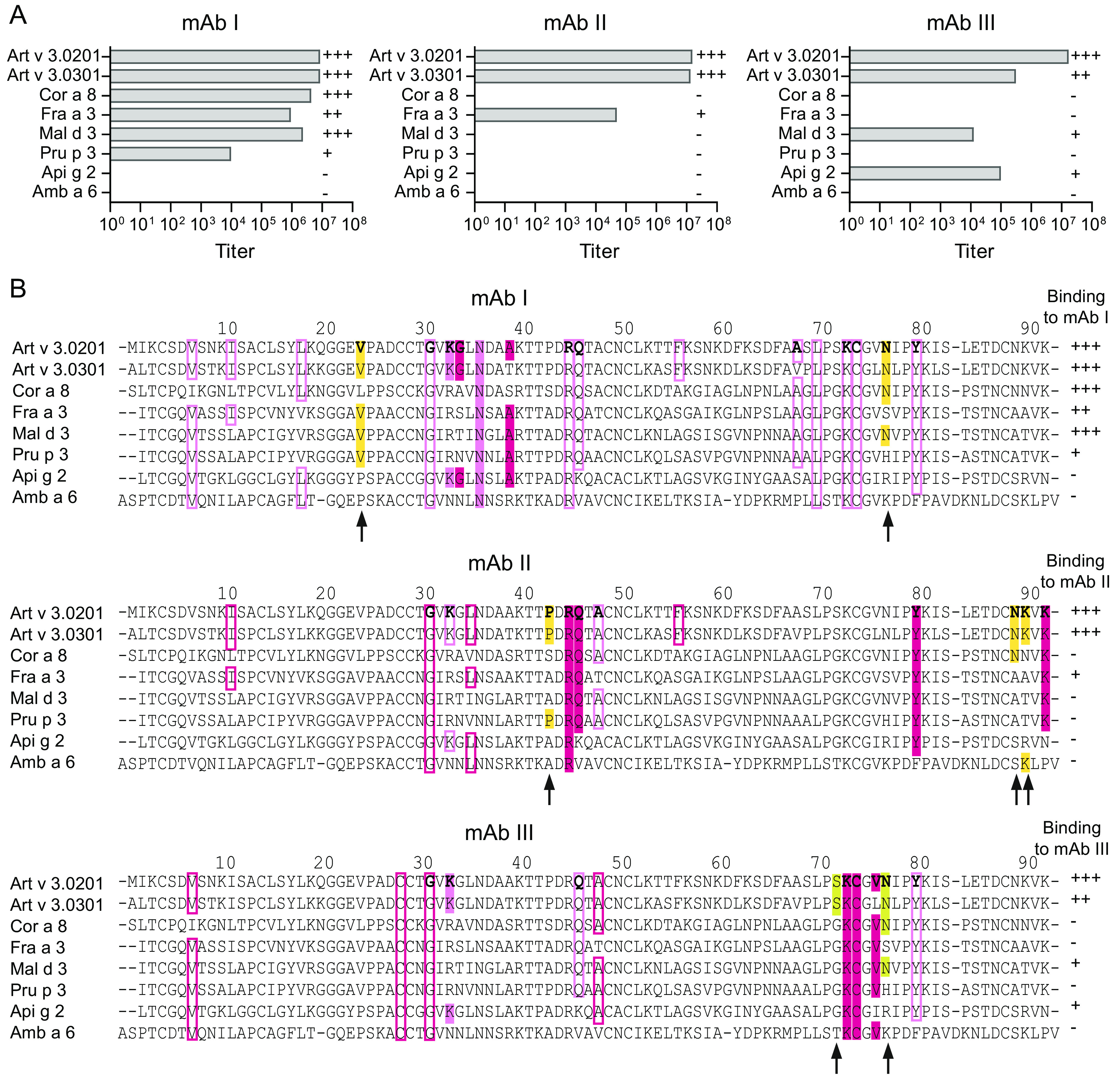
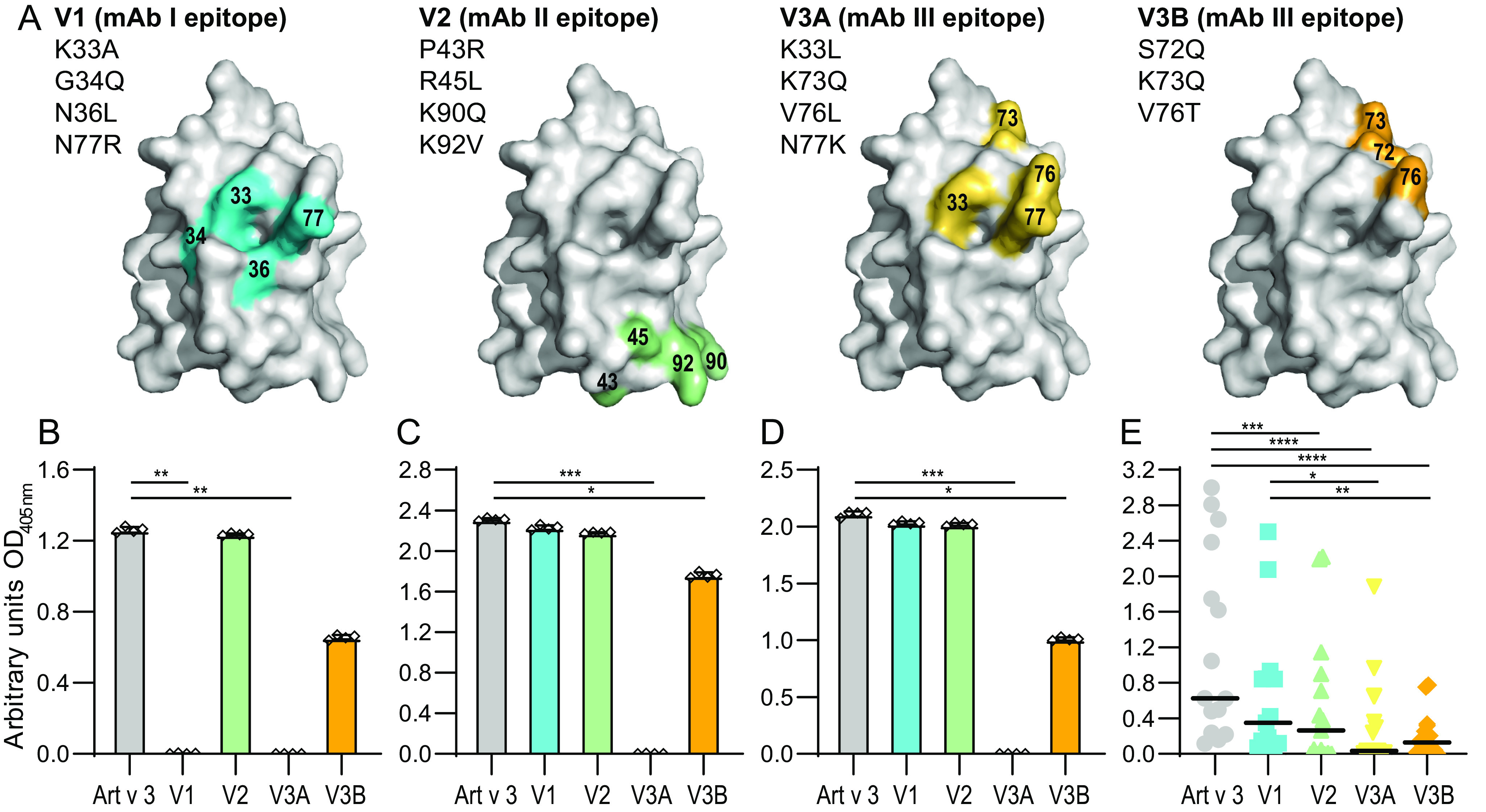
Identification of antibody-binding epitopes is crucial to understand immunological mechanisms. It is of particular interest for allergenic proteins with high cross-reactivity as observed in the lipid transfer protein (LTP) syndrome, which is characterized by severe allergic reactions. Art v 3, a pollen LTP from mugwort, is frequently involved in this cross-reactivity, but no antibody-binding epitopes have been determined so far. To reveal human IgE-binding regions of Art v 3, we produced three murine high-affinity mAbs, which showed 70-90% coverage of the allergenic epitopes from mugwort pollen-allergic patients. As reliable methods to determine structural epitopes with tightly interacting intact antibodies under native conditions are lacking, we developed a straightforward NMR approach termed hydrogen/deuterium exchange memory (HDXMEM). It relies on the slow exchange between the invisible antigen-mAb complex and the free 15N-labeled antigen whose 1H-15N correlations are detected. Due to a memory effect, changes of NH protection during antibody binding are measured. Differences in H/D exchange rates and analyses of mAb reactivity to homologous LTPs revealed three structural epitopes: two partially cross-reactive regions around α-helices 2 and 4 as well as a novel Art v 3-specific epitope at the C terminus. Protein variants with exchanged epitope residues confirmed the antibody-binding sites and revealed strongly reduced IgE reactivity. Using the novel HDXMEM for NMR epitope mapping allowed identification of the first structural epitopes of an allergenic pollen LTP. This knowledge enables improved cross-reactivity prediction for patients suffering from LTP allergy and facilitates design of therapeutics.
Keywords: Art v 3; NMR spectroscopy; allergen; allergen epitope mapping; epitope mapping; hydrogen/deuterium exchange; immunoglobulin E (IgE); immunoglobulin G (IgG); lipid transfer protein; monoclonal antibody; mugwort pollen; nuclear magnetic resonance (NMR).
Copyright © 2020 © 2020 Di Muzio et al. Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
R. v. R. reports consultancies from HAL Allergy BV, Citeq BV, and Amgany Inc. and speaker's fees from HAL Allergy BV, and Thermo Fisher Scientific. F. F. is a member of the Scientific Advisory Boards of HAL Allergy BV, AllergenOnline, and SIAF. G. G. is member of the Scientific Advisory Board of Bencard
Figures
References
-
- Mirza O., Henriksen A., Ipsen H., Larsen J. N., Wissenbach M., Spangfort M. D., and Gajhede M. (2000) Dominant epitopes and allergic cross-reactivity: complex formation between a Fab fragment of a monoclonal murine IgG antibody and the major allergen from birch pollen Bet v 1. J. Immunol. 165, 331–338 10.4049/jimmunol.165.1.331 - DOI - PubMed
-
- Padavattan S., Flicker S., Schirmer T., Madritsch C., Randow S., Reese G., Vieths S., Lupinek C., Ebner C., Valenta R., and Markovic-Housley Z. (2009) High-affinity IgE recognition of a conformational epitope of the major respiratory allergen Phl p 2 as revealed by X-ray crystallography. J. Immunol. 182, 2141–2151 10.4049/jimmunol.0803018 - DOI - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
