

N-acetylglucosamine drives myelination by triggering oligodendrocyte precursor cell differentiation
- PMID: 33453988
- PMCID: PMC7762951
- DOI: 10.1074/jbc.RA120.015595
N-acetylglucosamine drives myelination by triggering oligodendrocyte precursor cell differentiation
Abstract
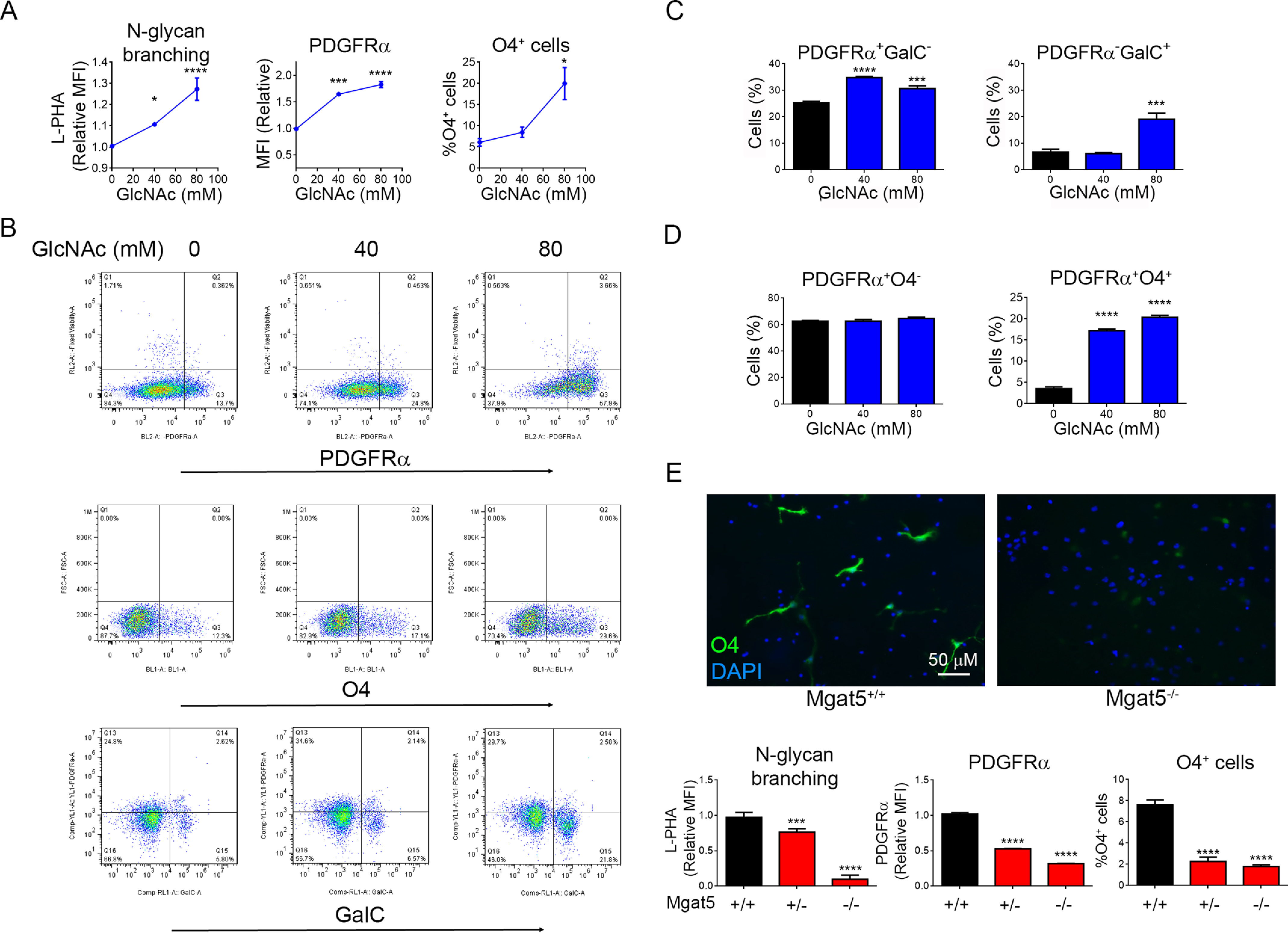
Myelination plays an important role in cognitive development and in demyelinating diseases like multiple sclerosis (MS), where failure of remyelination promotes permanent neuro-axonal damage. Modification of cell surface receptors with branched N-glycans coordinates cell growth and differentiation by controlling glycoprotein clustering, signaling, and endocytosis. GlcNAc is a rate-limiting metabolite for N-glycan branching. Here we report that GlcNAc and N-glycan branching trigger oligodendrogenesis from precursor cells by inhibiting platelet-derived growth factor receptor-α cell endocytosis. Supplying oral GlcNAc to lactating mice drives primary myelination in newborn pups via secretion in breast milk, whereas genetically blocking N-glycan branching markedly inhibits primary myelination. In adult mice with toxin (cuprizone)-induced demyelination, oral GlcNAc prevents neuro-axonal damage by driving myelin repair. In MS patients, endogenous serum GlcNAc levels inversely correlated with imaging measures of demyelination and microstructural damage. Our data identify N-glycan branching and GlcNAc as critical regulators of primary myelination and myelin repair and suggest that oral GlcNAc may be neuroprotective in demyelinating diseases like MS.
Keywords: N-acetylglucosamine; N-glycan branching; N-linked glycosylation; metabolism; multiple sclerosis; myelin; myelin repair; myelination; oligodendrocyte; oligodendrocyte precursor cell; oligodendrocytes.
Copyright © 2020 © 2020 Sy et al. Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
A. U. B., F. P., J. D., and M. D. are named as inventors on a patent that describes GlcNAc as a biomarker for multiple sclerosis. J. D. and M. D. are named as inventors on a patent for use of GlcNAc in multiple sclerosis and were cofounders of Glixis Therapeutics, a company developing analogs of GlcNAc for multiple sclerosis and other autoimmune diseases
Figures



References
-
- Lee Y., Morrison B. M., Li Y., Lengacher S., Farah M. H., Hoffman P. N., Liu Y., Tsingalia A., Jin L., Zhang P. W., Pellerin L., Magistretti P. J., and Rothstein J. D. (2012) Oligodendroglia metabolically support axons and contribute to neurodegeneration. Nature 487, 443–448 10.1038/nature11314 - DOI - PMC - PubMed
-
- Fünfschilling U., Supplie L. M., Mahad D., Boretius S., Saab A. S., Edgar J., Brinkmann B. G., Kassmann C. M., Tzvetanova I. D., Möbius W., Diaz F., Meijer D., Suter U., Hamprecht B., Sereda M. W., et al. (2012) Glycolytic oligodendrocytes maintain myelin and long-term axonal integrity. Nature 485, 517–521 10.1038/nature11007 - DOI - PMC - PubMed
-
- Mei F., Lehmann-Horn K., Shen Y.-A. A., Rankin K. A., Stebbins K. J., Lorrain D. S., Pekarek K., A Sagan S., Xiao L., Teuscher C., von Büdingen H.-C., Wess J., Lawrence J. J., Green A. J., Fancy S. P., et al. (2016) Accelerated remyelination during inflammatory demyelination prevents axonal loss and improves functional recovery. eLife 5, 10.7554/eLife.18246 - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
