

Evolving the naturally compromised chorismate mutase from Mycobacterium tuberculosis to top performance
- PMID: 33453995
- PMCID: PMC7762937
- DOI: 10.1074/jbc.RA120.014924
Evolving the naturally compromised chorismate mutase from Mycobacterium tuberculosis to top performance
Abstract
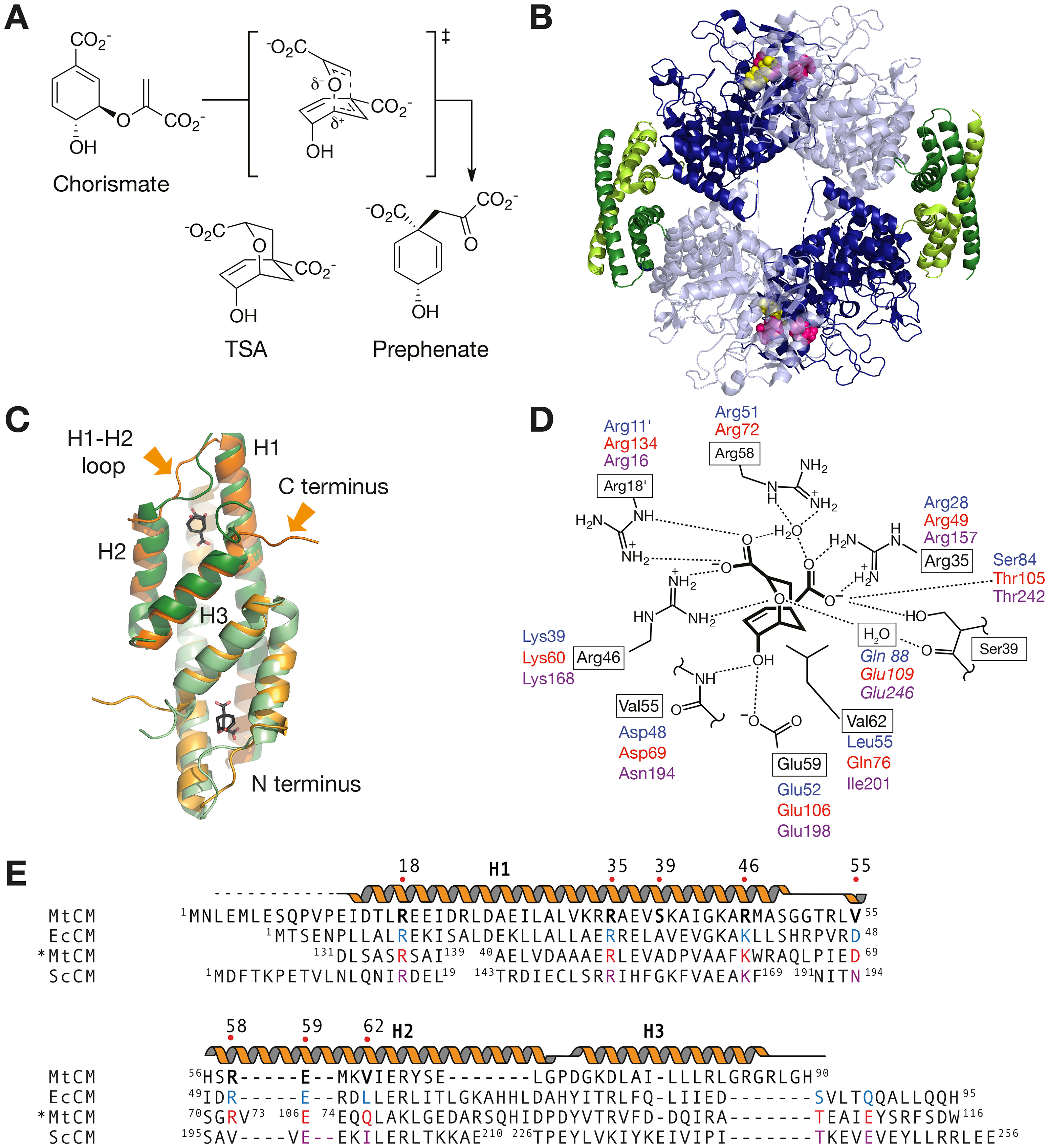
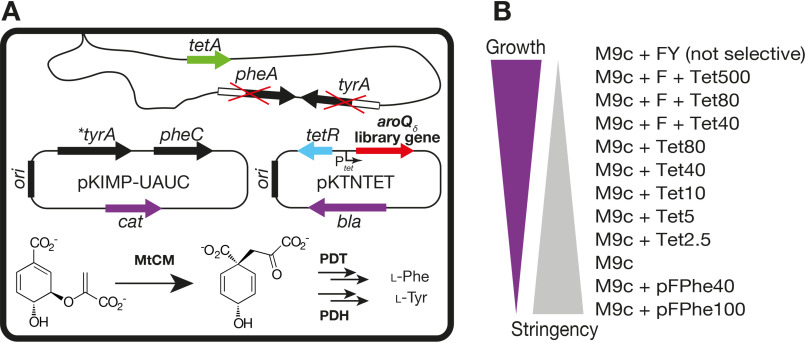
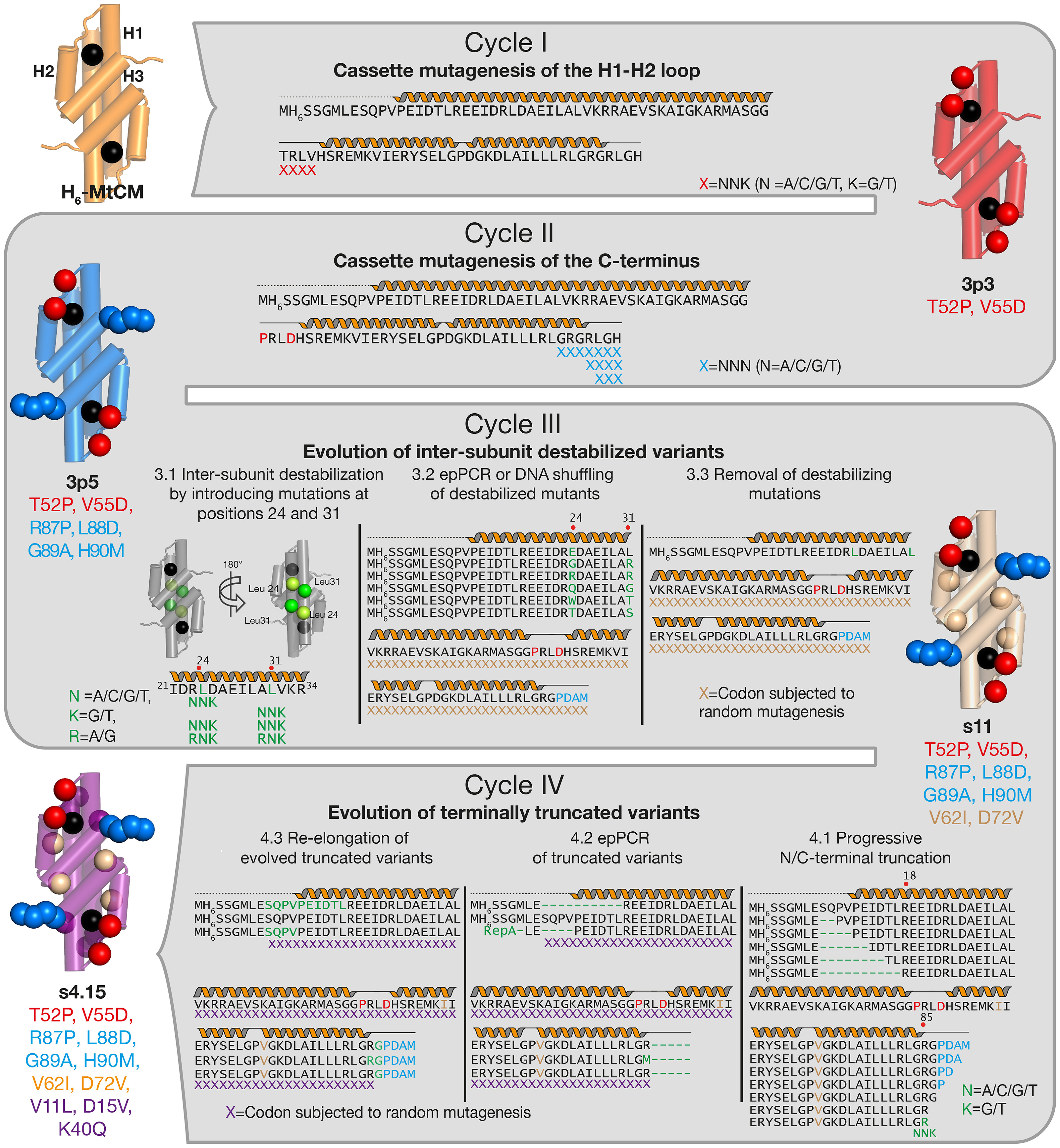
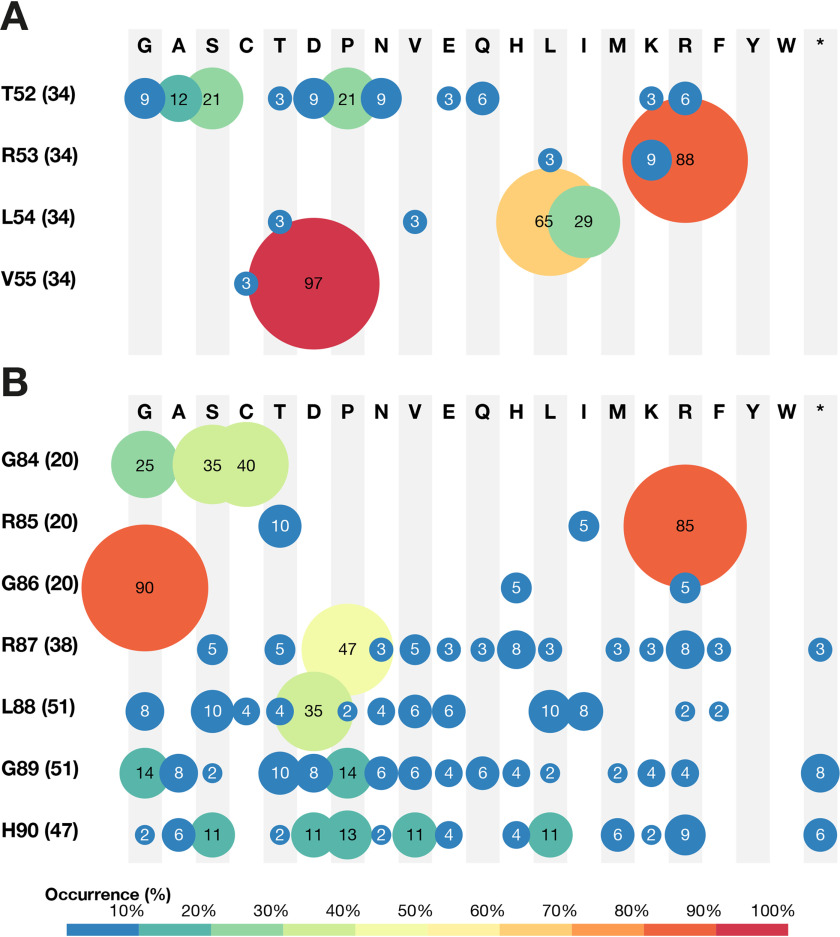
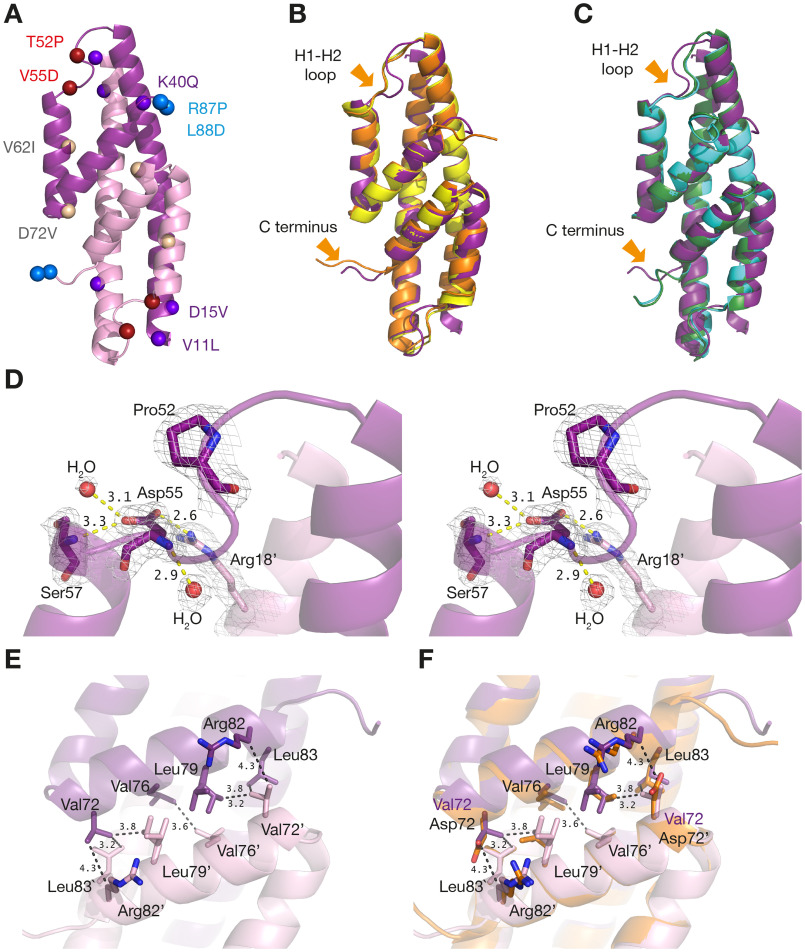
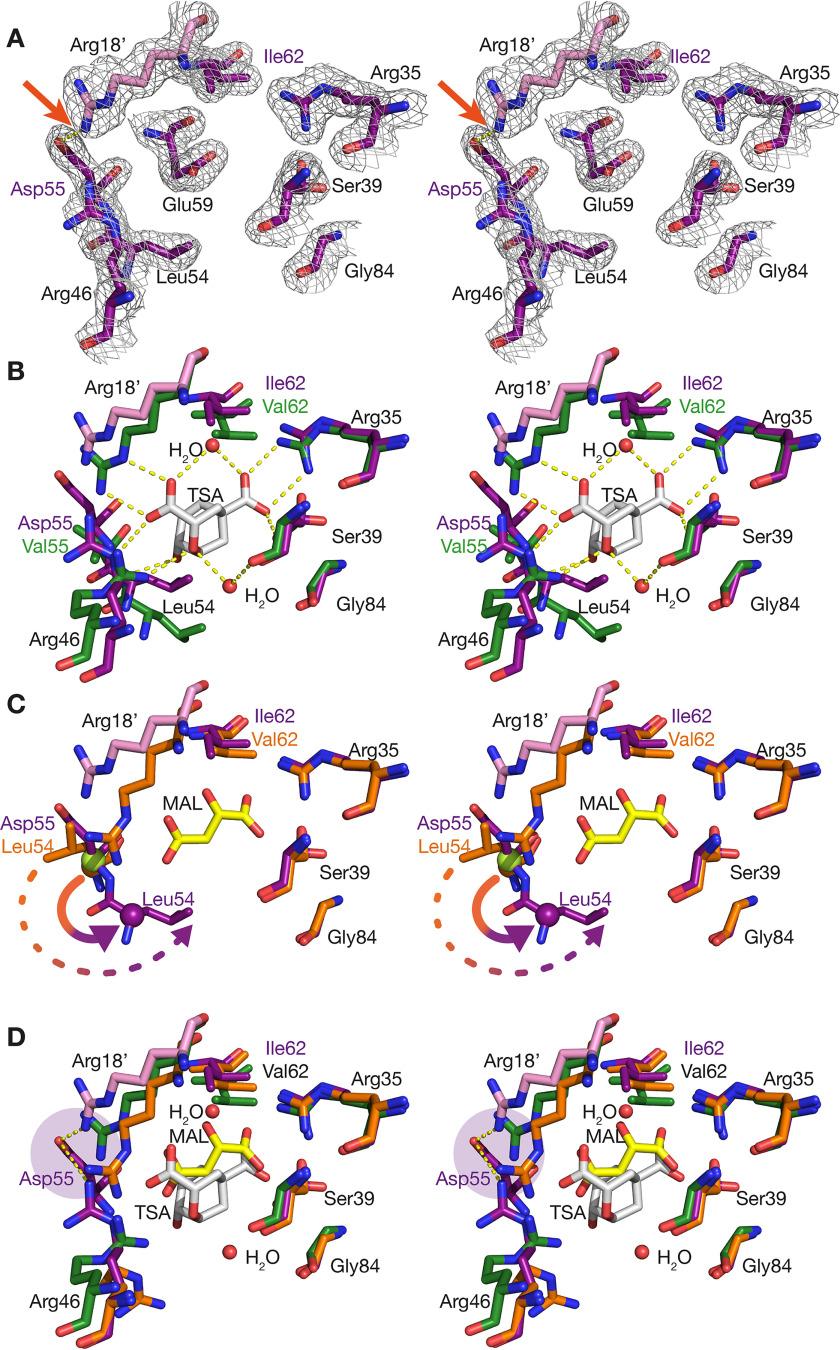
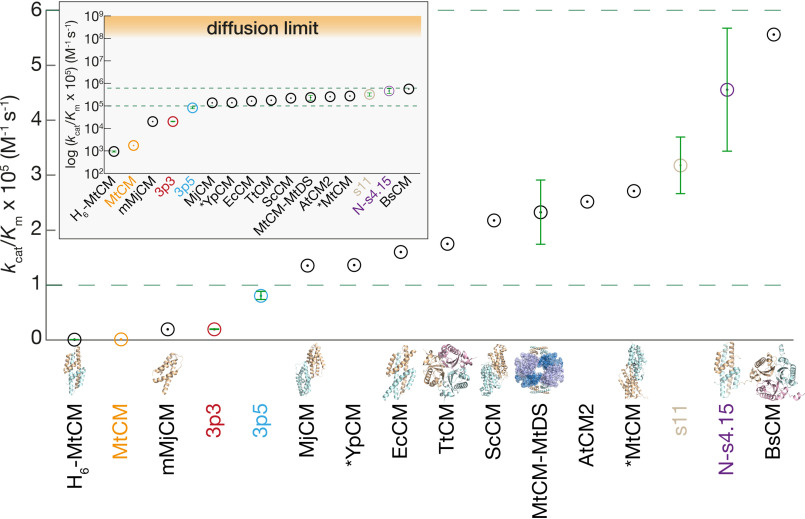
Chorismate mutase (CM), an essential enzyme at the branch-point of the shikimate pathway, is required for the biosynthesis of phenylalanine and tyrosine in bacteria, archaea, plants, and fungi. MtCM, the CM from Mycobacterium tuberculosis, has less than 1% of the catalytic efficiency of a typical natural CM and requires complex formation with 3-deoxy-d-arabino-heptulosonate 7-phosphate synthase for high activity. To explore the full potential of MtCM for catalyzing its native reaction, we applied diverse iterative cycles of mutagenesis and selection, thereby raising kcat/Km 270-fold to 5 × 105m-1s-1, which is even higher than for the complex. Moreover, the evolutionarily optimized autonomous MtCM, which had 11 of its 90 amino acids exchanged, was stabilized compared with its progenitor, as indicated by a 9 °C increase in melting temperature. The 1.5 Å crystal structure of the top-evolved MtCM variant reveals the molecular underpinnings of this activity boost. Some acquired residues (e.g. Pro52 and Asp55) are conserved in naturally efficient CMs, but most of them lie beyond the active site. Our evolutionary trajectories reached a plateau at the level of the best natural enzymes, suggesting that we have exhausted the potential of MtCM. Taken together, these findings show that the scaffold of MtCM, which naturally evolved for mediocrity to enable inter-enzyme allosteric regulation of the shikimate pathway, is inherently capable of high activity.
Keywords: X-ray crystallography; catalytic efficiency; conformational change; crystal structure; directed evolution; enzyme catalysis; enzyme mutation; molecular evolution; protein structure; structure-activity relationship.
Copyright © 2020 © 2020 Fahrig-Kamarauskait? et al. Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
The authors declare that they have no conflicts of interest with the contents of this article
Figures
Similar articles
-
Structure and function of a complex between chorismate mutase and DAHP synthase: efficiency boost for the junior partner.EMBO J. 2009 Jul 22;28(14):2128-42. doi: 10.1038/emboj.2009.165. Epub 2009 Jun 25. EMBO J. 2009. PMID: 19556970 Free PMC article.
-
Remote Control by Inter-Enzyme Allostery: A Novel Paradigm for Regulation of the Shikimate Pathway.J Mol Biol. 2016 Mar 27;428(6):1237-1255. doi: 10.1016/j.jmb.2016.01.001. Epub 2016 Jan 8. J Mol Biol. 2016. PMID: 26776476
-
A novel noncovalent complex of chorismate mutase and DAHP synthase from Mycobacterium tuberculosis: protein purification, crystallization and X-ray diffraction analysis.Acta Crystallogr Sect F Struct Biol Cryst Commun. 2009 Oct 1;65(Pt 10):1048-52. doi: 10.1107/S1744309109035878. Epub 2009 Sep 25. Acta Crystallogr Sect F Struct Biol Cryst Commun. 2009. PMID: 19851019 Free PMC article.
-
Mycobacterium tuberculosis chorismate mutase: A potential target for TB.Bioorg Med Chem. 2017 Mar 15;25(6):1725-1736. doi: 10.1016/j.bmc.2017.02.001. Epub 2017 Feb 4. Bioorg Med Chem. 2017. PMID: 28202315 Review.
-
Pericyclic reactions catalyzed by chorismate-utilizing enzymes.Biochemistry. 2011 Sep 6;50(35):7476-83. doi: 10.1021/bi2009739. Epub 2011 Aug 12. Biochemistry. 2011. PMID: 21823653 Free PMC article. Review.
Cited by
-
What Drives Chorismate Mutase to Top Performance? Insights from a Combined In Silico and In Vitro Study.Biochemistry. 2023 Feb 7;62(3):782-796. doi: 10.1021/acs.biochem.2c00635. Epub 2023 Jan 27. Biochemistry. 2023. PMID: 36705397 Free PMC article.
-
A Robust Growth-Based Selection Platform to Evolve an Enzyme via Dependency on Noncanonical Tyrosine Analogues.JACS Au. 2024 Mar 19;4(4):1583-1590. doi: 10.1021/jacsau.4c00070. eCollection 2024 Apr 22. JACS Au. 2024. PMID: 38665651 Free PMC article.
-
Adapting enzymes to improve their functionality in plants: why and how.Biochem Soc Trans. 2023 Oct 31;51(5):1957-1966. doi: 10.1042/BST20230532. Biochem Soc Trans. 2023. PMID: 37787016 Free PMC article. Review.
References
-
- Burschowsky D., Thorbjørnsrud H. V., Heim J. B., Fahrig-Kamarauskaitė J., Würth-Roderer K., Kast P., and Krengel U. (2018) Inter-enzyme allosteric regulation of chorismate mutase in Corynebacterium glutamicum: structural basis of feedback activation by Trp. Biochemistry 57, 557–573 10.1021/acs.biochem.7b01018 - DOI - PubMed
-
- Haslam E. (1993) Shikimic Acid: Metabolism and Metabolites, John Wiley, New York
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous
