

Bacterial iron detoxification at the molecular level
- PMID: 33454001
- PMCID: PMC7762939
- DOI: 10.1074/jbc.REV120.007746
Bacterial iron detoxification at the molecular level
Abstract
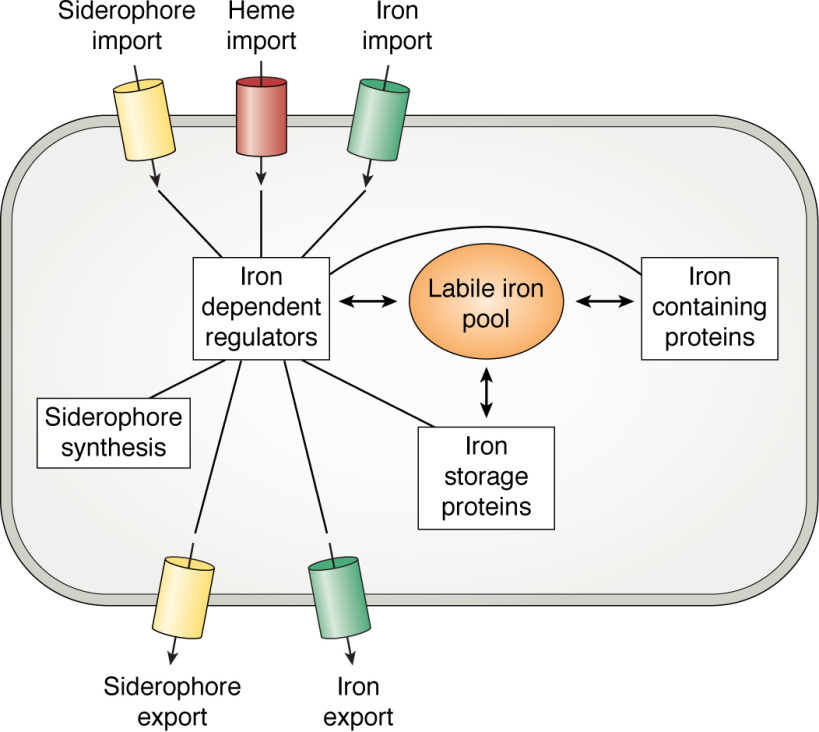
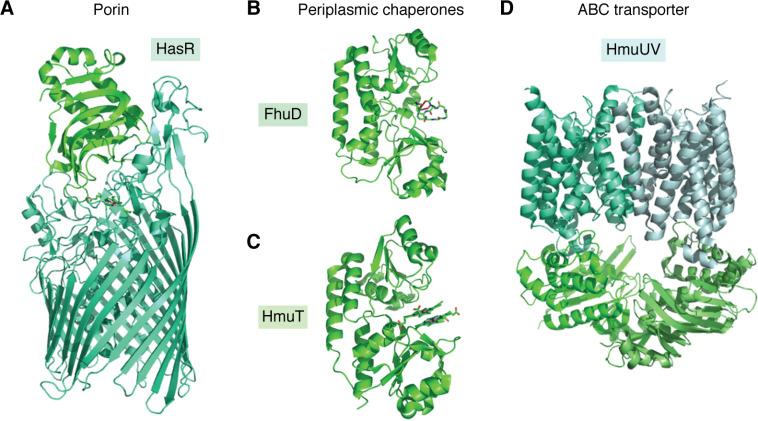
Iron is an essential micronutrient, and, in the case of bacteria, its availability is commonly a growth-limiting factor. However, correct functioning of cells requires that the labile pool of chelatable "free" iron be tightly regulated. Correct metalation of proteins requiring iron as a cofactor demands that such a readily accessible source of iron exist, but overaccumulation results in an oxidative burden that, if unchecked, would lead to cell death. The toxicity of iron stems from its potential to catalyze formation of reactive oxygen species that, in addition to causing damage to biological molecules, can also lead to the formation of reactive nitrogen species. To avoid iron-mediated oxidative stress, bacteria utilize iron-dependent global regulators to sense the iron status of the cell and regulate the expression of proteins involved in the acquisition, storage, and efflux of iron accordingly. Here, we survey the current understanding of the structure and mechanism of the important members of each of these classes of protein. Diversity in the details of iron homeostasis mechanisms reflect the differing nutritional stresses resulting from the wide variety of ecological niches that bacteria inhabit. However, in this review, we seek to highlight the similarities of iron homeostasis between different bacteria, while acknowledging important variations. In this way, we hope to illustrate how bacteria have evolved common approaches to overcome the dual problems of the insolubility and potential toxicity of iron.
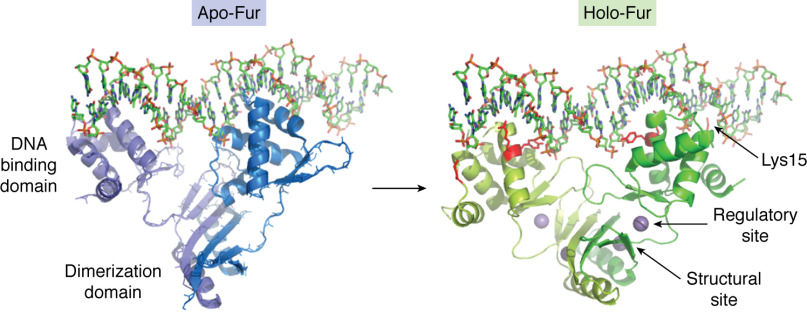
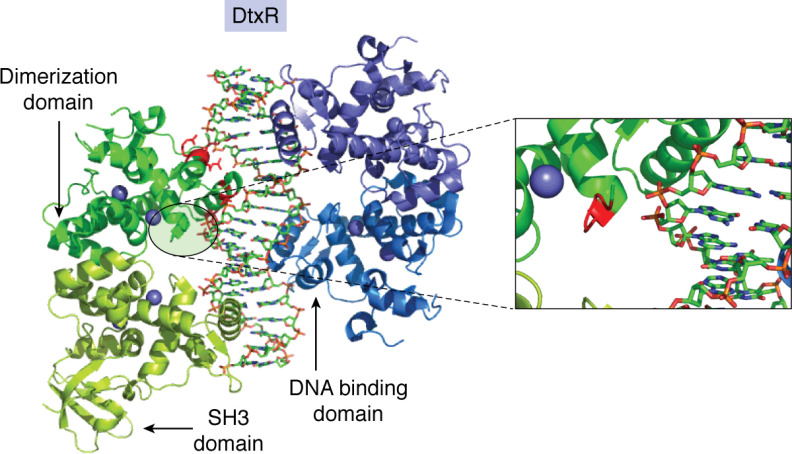
Keywords: Bfr; Dps; DtxR; Ftn; Fur; Irr; RirA; bacterioferritin; encapsulin; encapsulins; ferritin; gene regulation; iron; iron metabolism; iron regulation; iron storage; iron toxicity; reactive oxygen species (ROS).
Copyright © 2020 © 2020 Bradley et al. Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
The authors declare that they have no conflicts of interest with the contents of this article
Figures

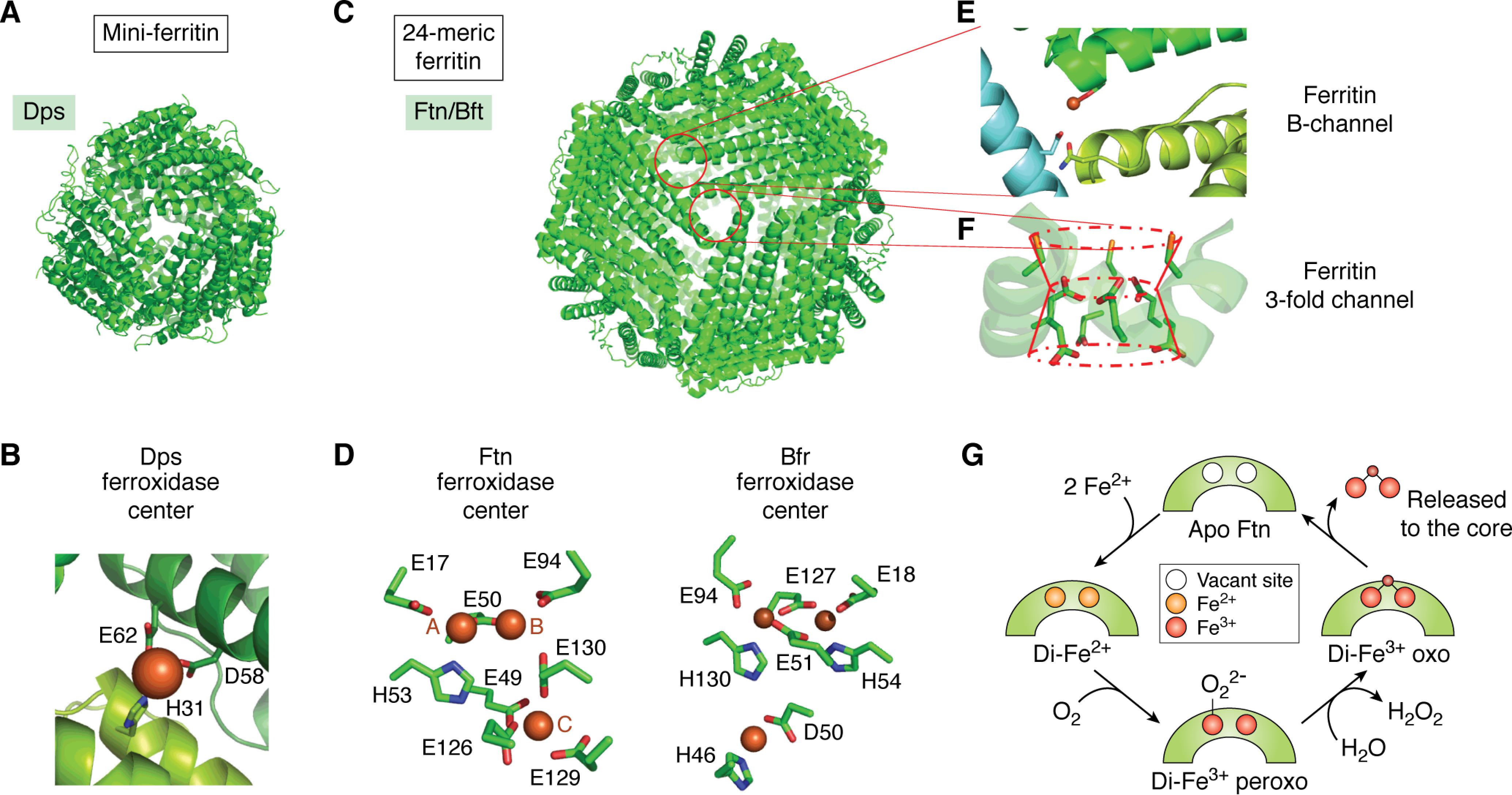
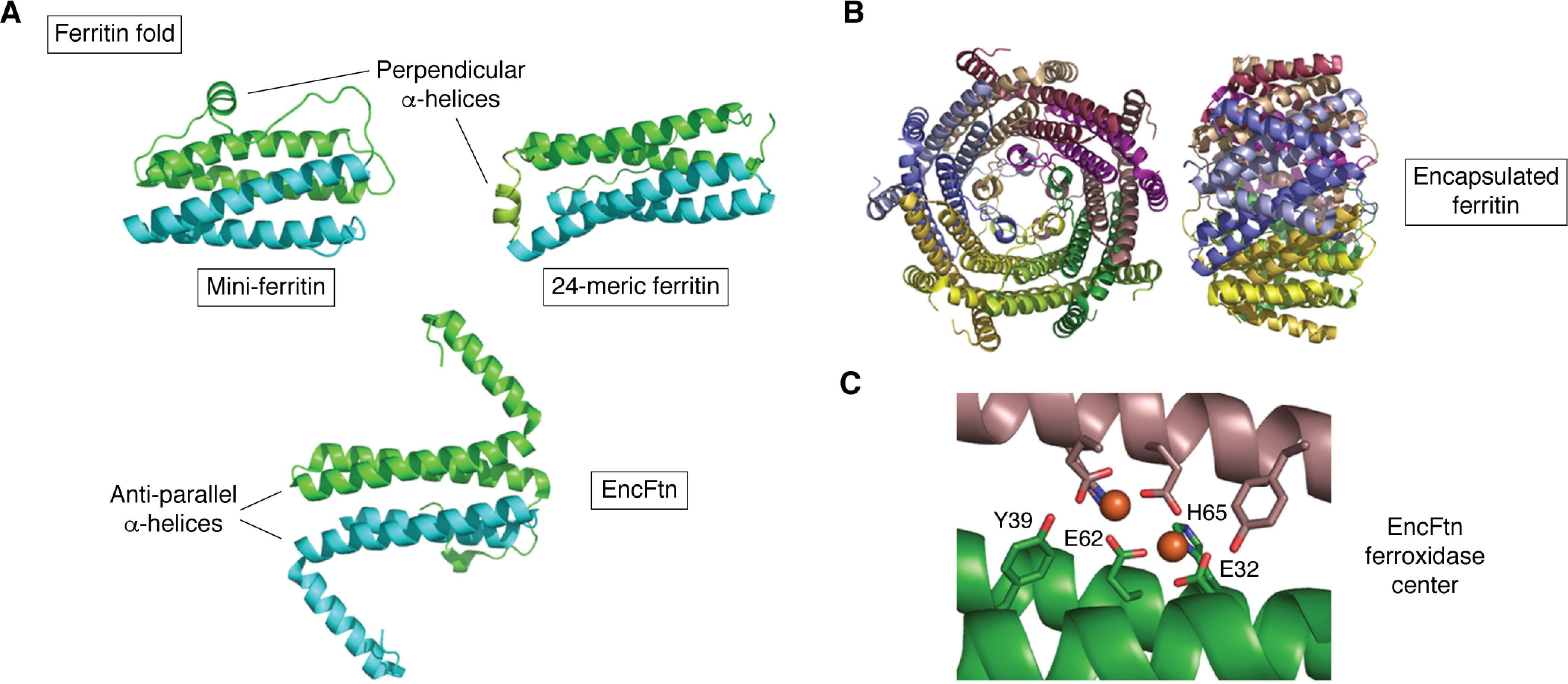
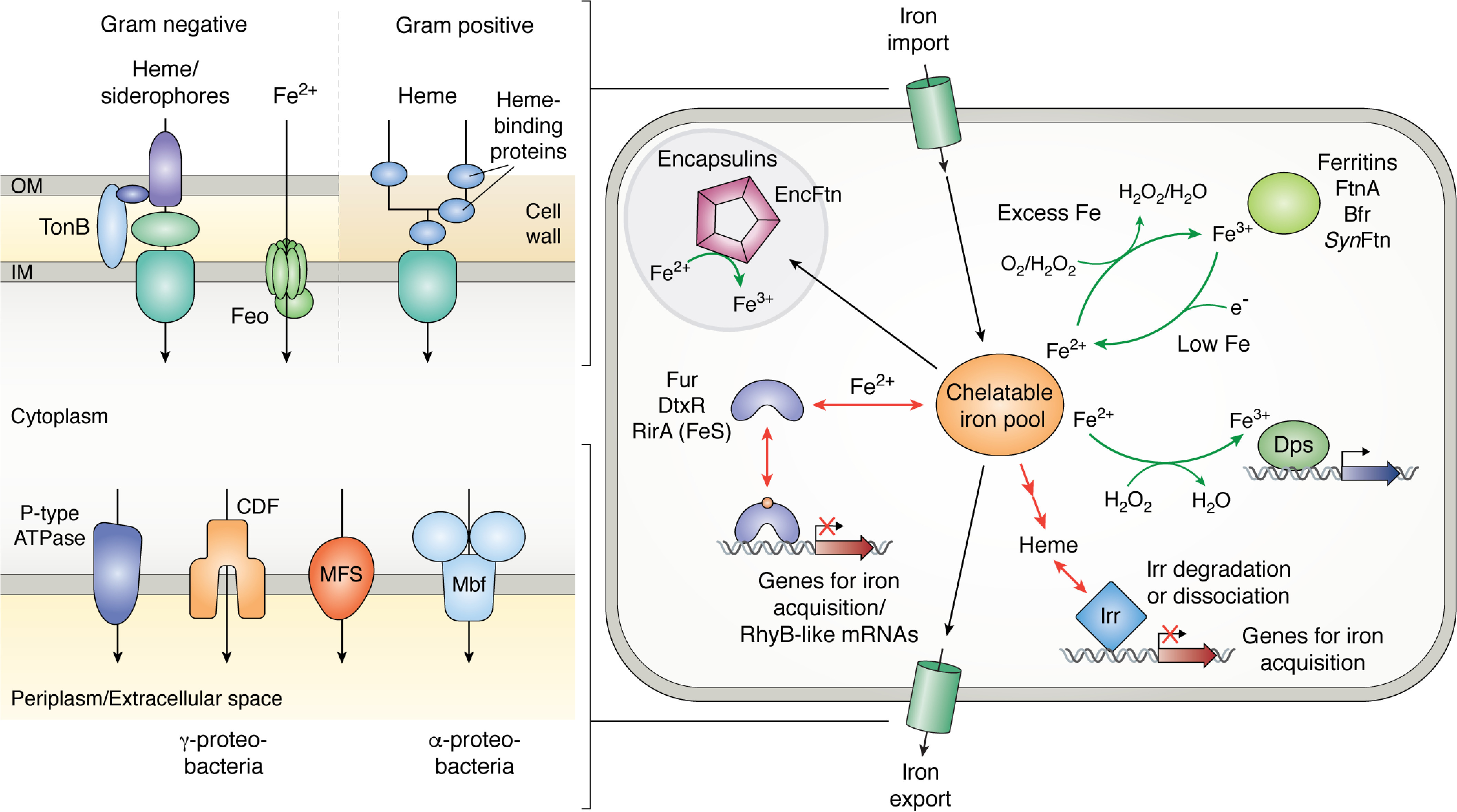
References
-
- Gamba I., Codolà Z., Lloret-Fillol J., and Costas M. (2017) Making and breaking of the O-O bond at iron complexes. Coord. Chem. Rev. 334, 2–24 10.1016/j.ccr.2016.11.007 - DOI
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
