

Anti-NKG2D single domain-based antibodies for the modulation of anti-tumor immune response
- PMID: 33457075
- PMCID: PMC7781768
- DOI: 10.1080/2162402X.2020.1854529
Anti-NKG2D single domain-based antibodies for the modulation of anti-tumor immune response
Abstract
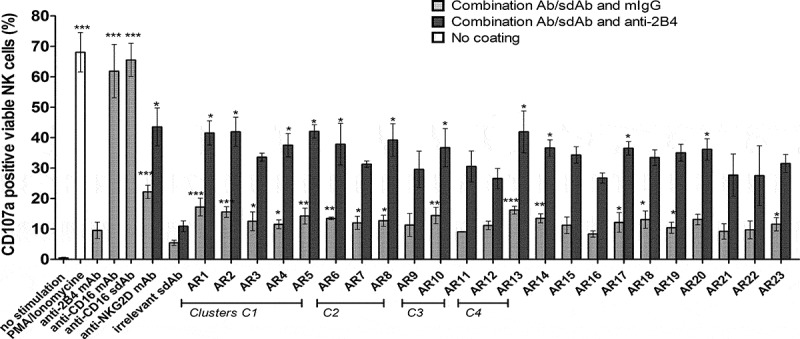
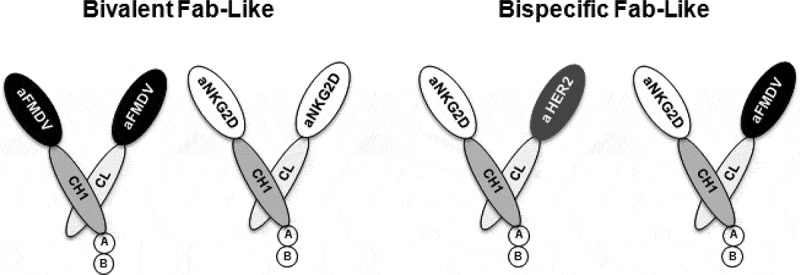
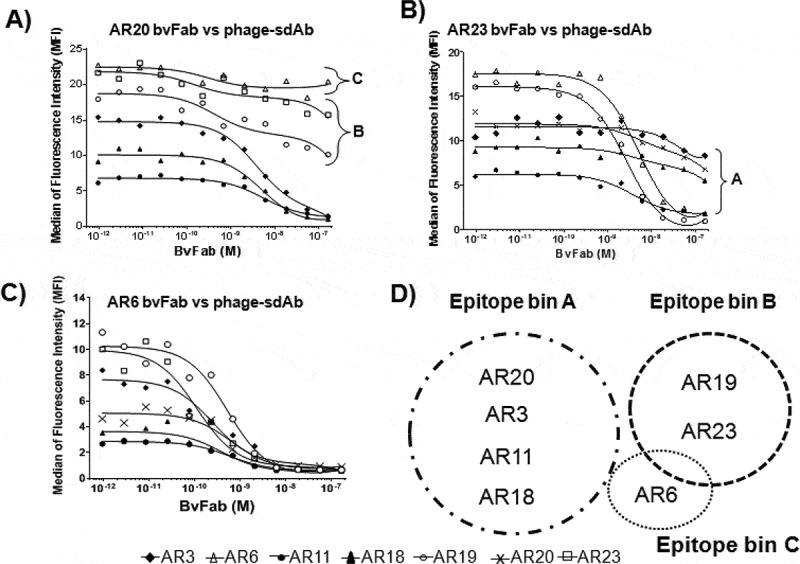
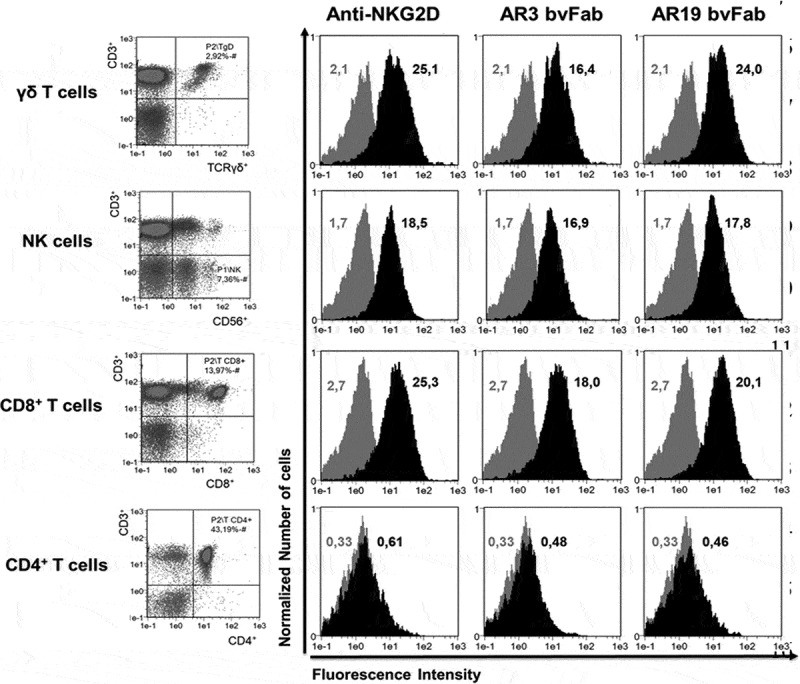
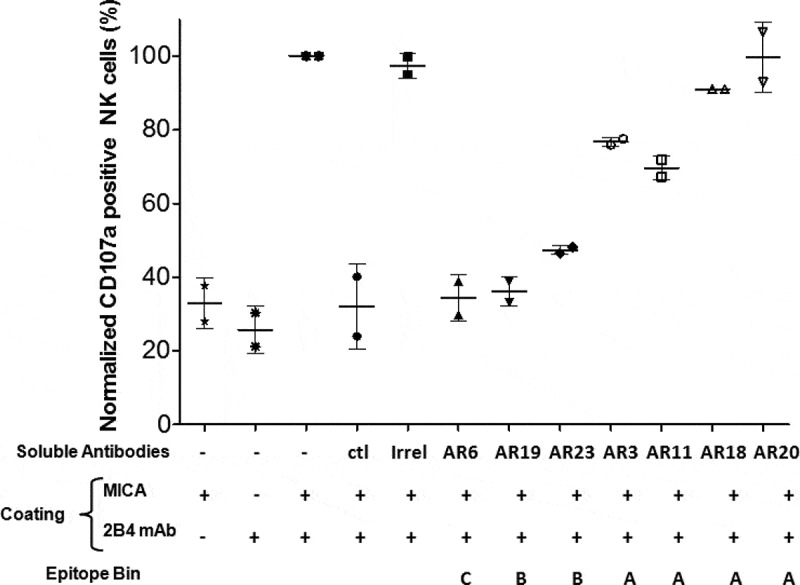
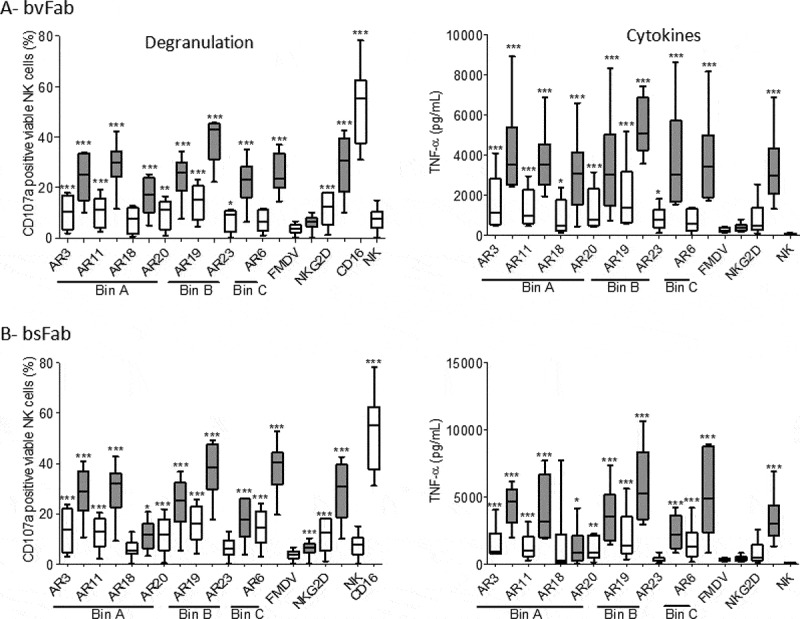
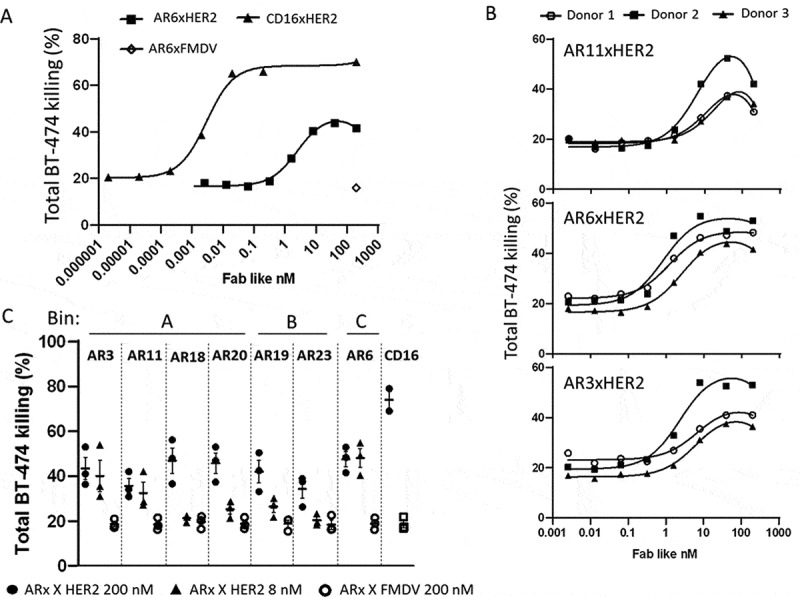
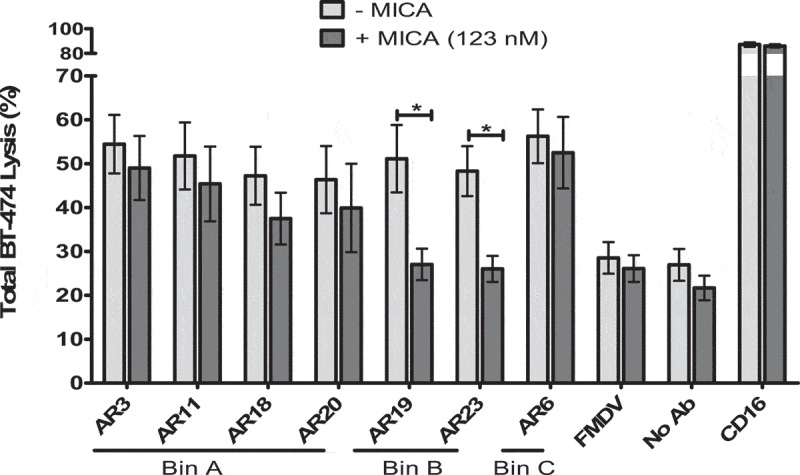
The natural killer group 2 member D (NKG2D) receptor is a C-type lectin-like activating receptor mainly expressed by cytotoxic immune cells including NK, CD8+ T, γδ T and NKT cells and in some pathological conditions by a subset of CD4+ T cells. It binds a variety of ligands (NKG2DL) whose expressions is finely regulated by stress-related conditions. The NKG2DL/NKG2D axis plays a central and complex role in the regulation of immune responses against diverse cellular threats such as oncogene-mediated transformations or infections. We generated a panel of seven highly specific anti-human NKG2D single-domain antibodies targeting various epitopes. These single-domain antibodies were integrated into bivalent and bispecific antibodies using a versatile plug-and-play Fab-like format. Depending on the context, these Fab-like antibodies exhibited activating or inhibitory effects on the immune response mediated by the NKG2DL/NKG2D axis. In solution, the bivalent anti-NKG2D antibodies that compete with NKG2DL potently blocked the activation of NK cells seeded on immobilized MICA, thus constituting antagonizing candidates. Bispecific anti-NKG2DxHER2 antibodies that concomitantly engage HER2 on tumor cells and NKG2D on NK cells elicited cytotoxicity of unstimulated NK in a tumor-specific manner, regardless of their apparent affinities and epitopes. Importantly, the bispecific antibodies that do not compete with ligands binding retained their full cytotoxic activity in the presence of ligands, a valuable property to circumvent immunosuppressive effects induced by soluble ligands in the microenvironment.
Keywords: HER2; NK cells; NKG2D/NKG2DL axis; cell engagers; single-domain antibody.
© 2020 The Author(s). Published with license by Taylor & Francis Group, LLC.
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
Miscellaneous