

A Tumor Suppressor Enhancer of PTEN in T-cell development and leukemia
- PMID: 33458694
- PMCID: PMC7810363
- DOI: 10.1158/2643-3230.BCD-20-0201
A Tumor Suppressor Enhancer of PTEN in T-cell development and leukemia
Abstract
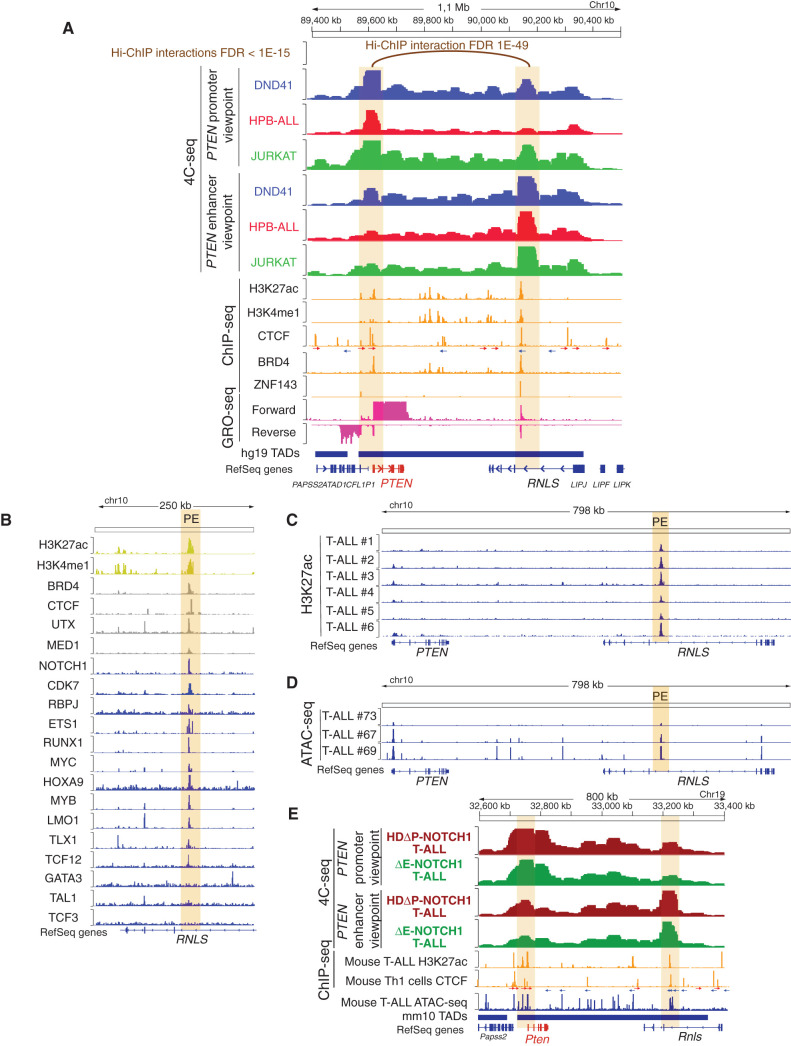
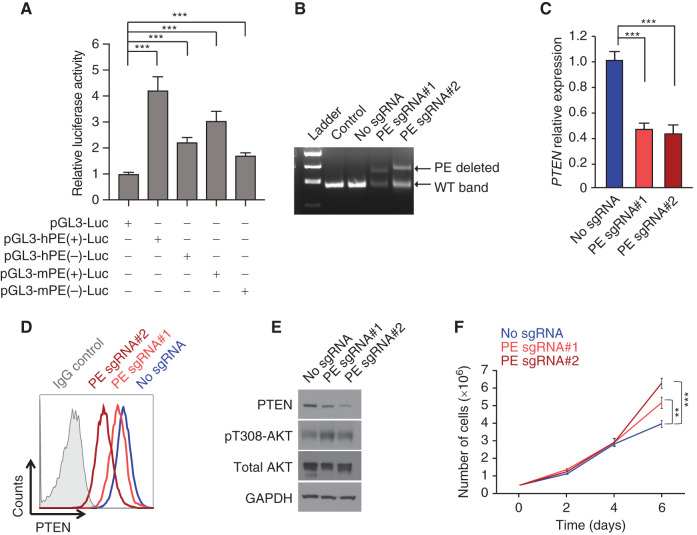
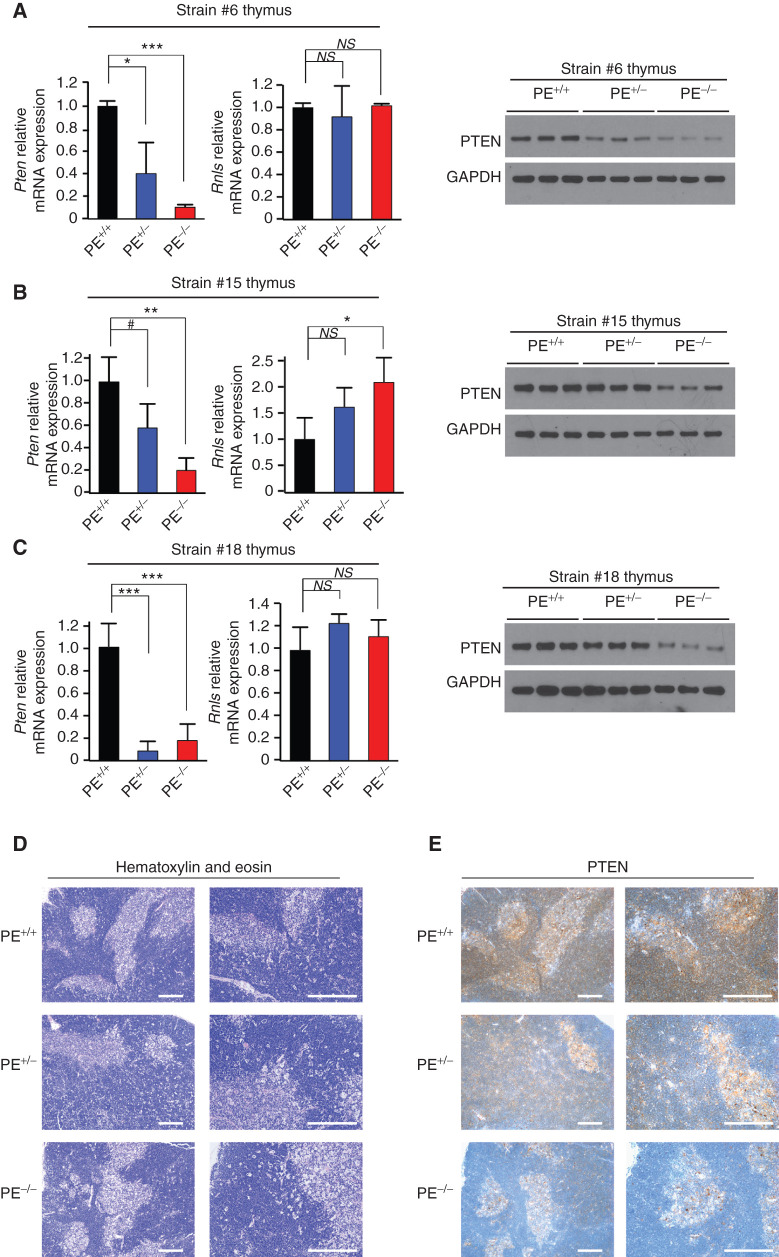
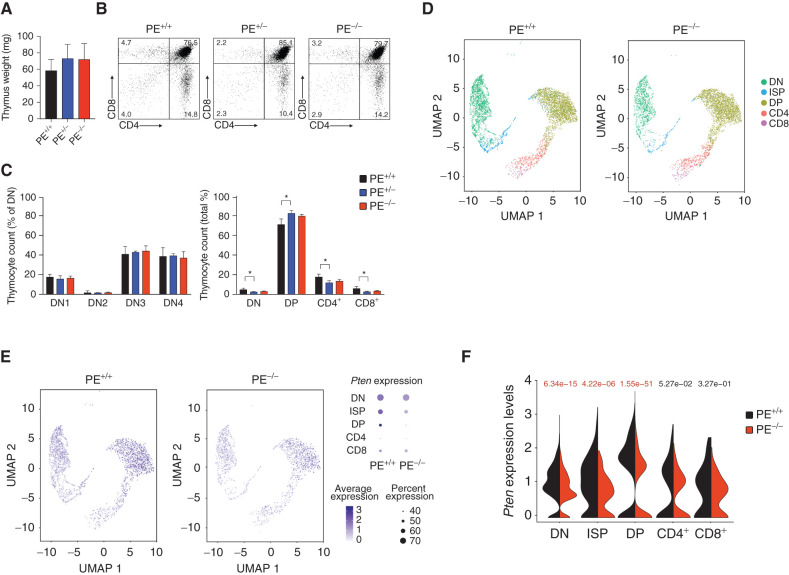
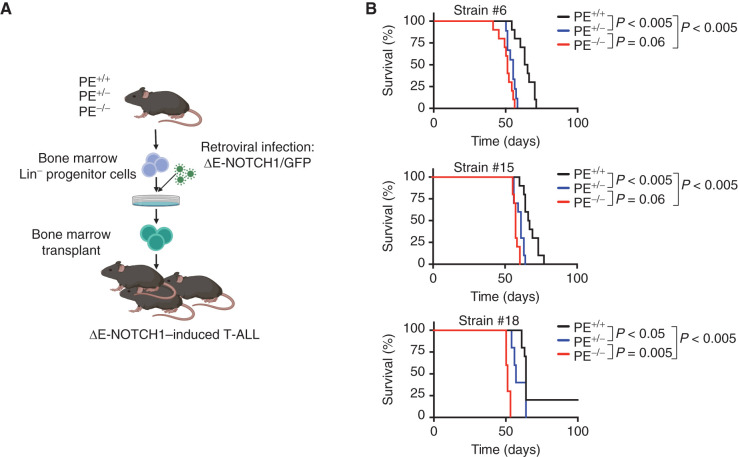
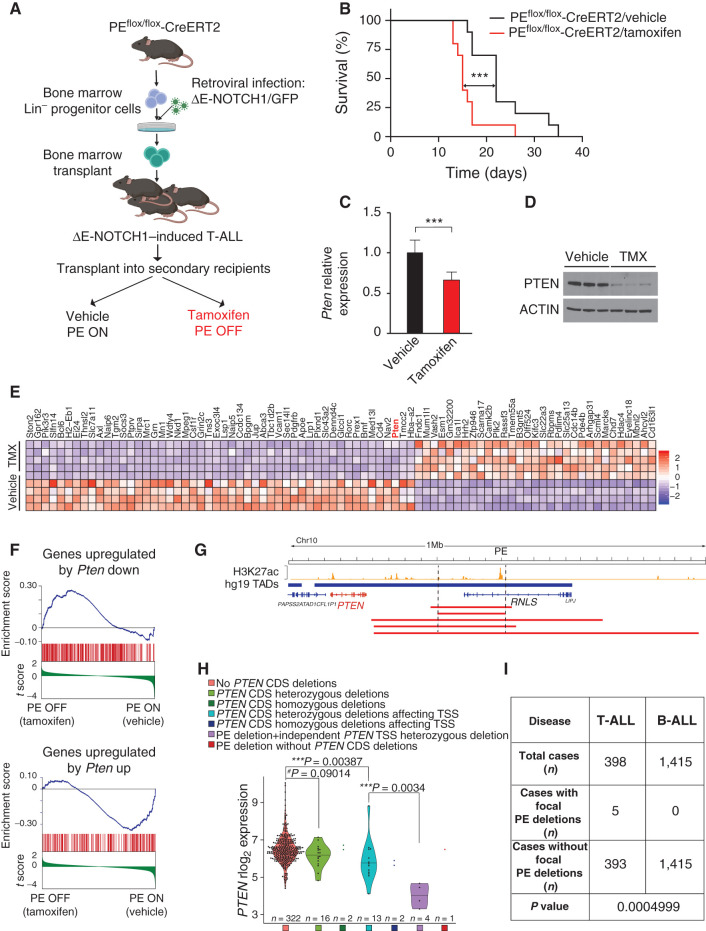
Long-range oncogenic enhancers play an important role in cancer. Yet, whether similar regulation of tumor suppressor genes is relevant remains unclear. Loss of expression of PTEN is associated with the pathogenesis of various cancers, including T-cell leukemia (T-ALL). Here, we identify a highly conserved distal enhancer (PE) that interacts with the PTEN promoter in multiple hematopoietic populations, including T-cells, and acts as a hub of relevant transcription factors in T-ALL. Consistently, loss of PE leads to reduced PTEN levels in T-ALL cells. Moreover, PE-null mice show reduced Pten levels in thymocytes and accelerated development of NOTCH1-induced T-ALL. Furthermore, secondary loss of PE in established leukemias leads to accelerated progression and a gene expression signature driven by Pten loss. Finally, we uncovered recurrent deletions encompassing PE in T-ALL, which are associated with decreased PTEN levels. Altogether, our results identify PE as the first long-range tumor suppressor enhancer directly implicated in cancer.
Keywords: NOTCH1; PTEN; T-ALL; T-cell acute lymphoblastic leukemia; enhancer.
Conflict of interest statement
Conflict of interest disclosure: The authors declare no competing financial interests.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
