

Measuring strain in the exoskeleton of spiders-virtues and caveats
- PMID: 33459819
- PMCID: PMC8046692
- DOI: 10.1007/s00359-020-01458-y
Measuring strain in the exoskeleton of spiders-virtues and caveats
Abstract
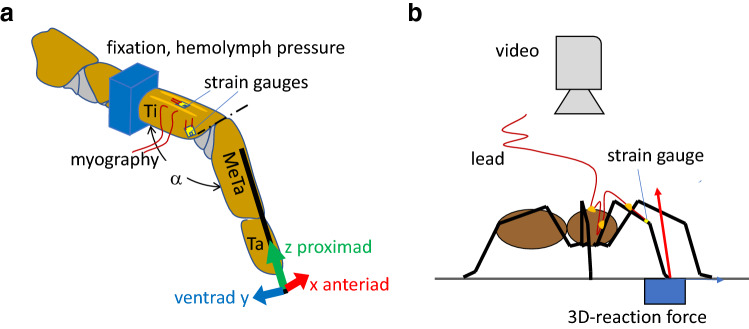
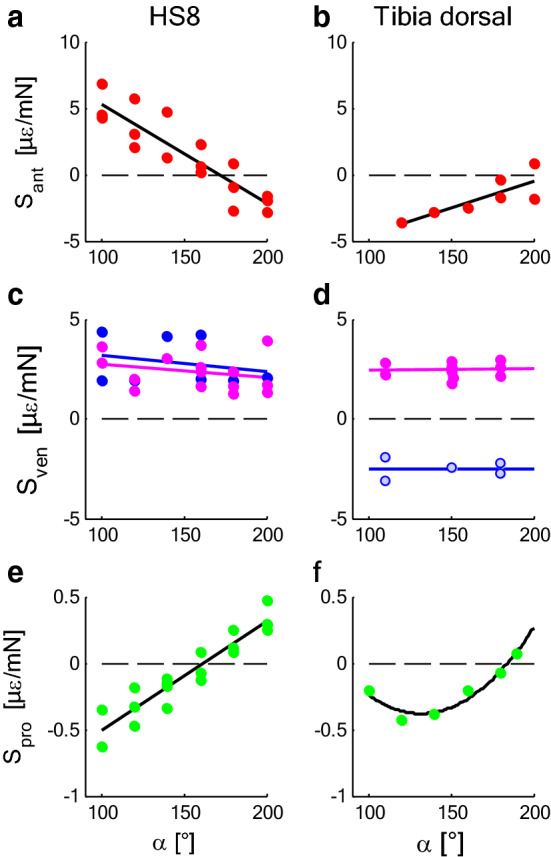
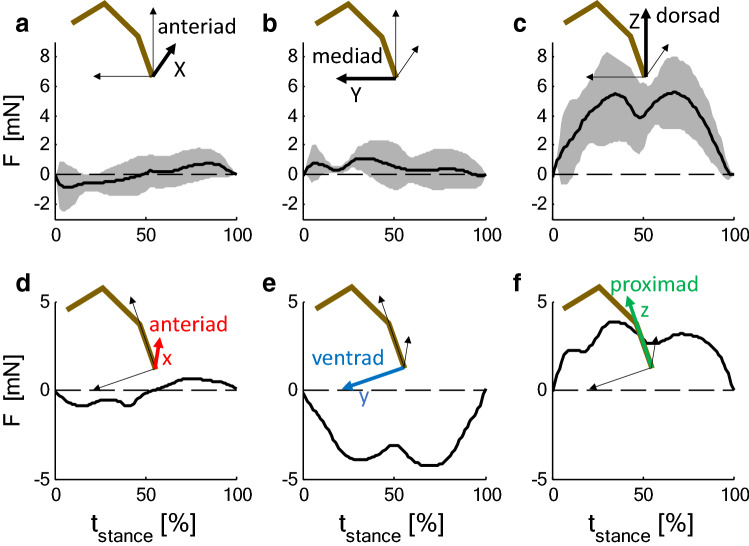
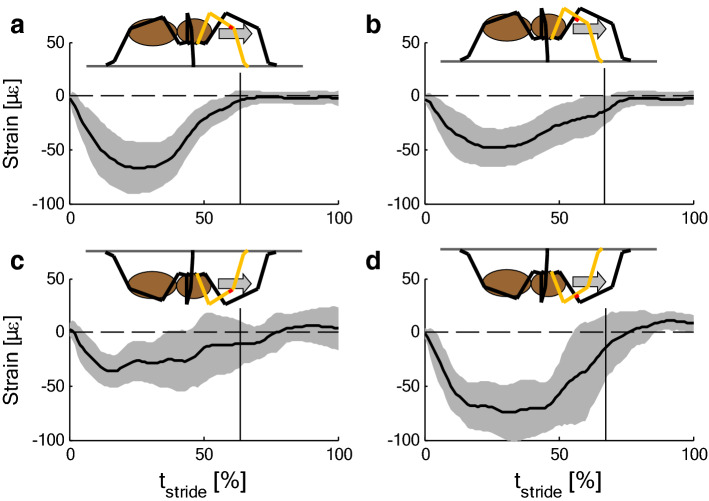
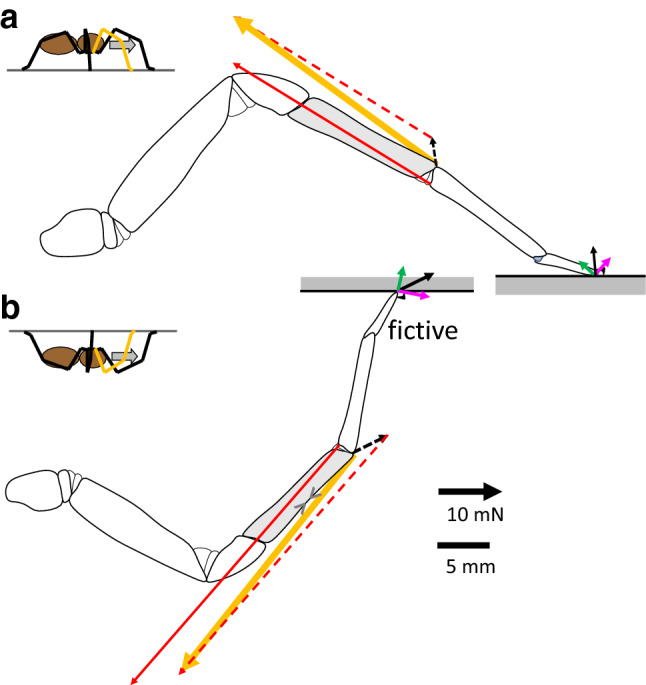
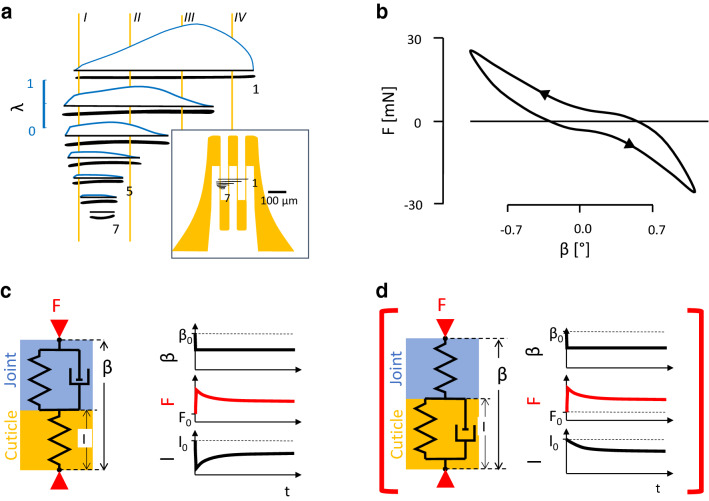
The measurement of cuticular strain during locomotion using foil strain gauges provides information both on the loads of the exoskeleton bears and the adaptive value of the specific location of natural strain detectors (slit sense organs). Here, we critically review available literature. In tethered animals, by applying loads to the metatarsus tip, strain and mechanical sensitivity (S = strain/load) induced at various sites in the tibia were determined. The loci of the lyriform organs close to the tibia-metatarsus joint did not stand out by high strain. The strains induced at various sites during free locomotion can be interpreted based on S and, beyond the joint region, on beam theory. Spiders avoided laterad loading of the tibia-metatarsus joint during slow locomotion. Balancing body weight, joint flexors caused compressive strain at the posterior and dorsal tibia. While climbing upside down strain measurements indicate strong flexor activity. In future studies, a precise calculation and quantitative determination of strain at the sites of the lyriform organs will profit from more detailed data on the overall strain distribution, morphology, and material properties. The values and caveats of the strain gauge technology, the only one applicable to freely moving spiders, are discussed.
Keywords: Joint load; Locomotion; Mechanical sensitivity; Sensory strain reception; Strain gauge.
Conflict of interest statement
No conflicts of interest.
Figures
References
-
- Barth F (eds) (2012) Learning from animal sensors: the clever “design” of spider mechanoreceptors. In: Bioinspiration, biomimetics, and bioreplication, vol 833904, SPIE, San Diego. 10.1117/12.902456
-
- Barth FG. A spider’s world: senses and behavior. Berlin: Springer; 2002.
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources
