

More than just protein building blocks: how amino acids and related metabolic pathways fuel macrophage polarization
- PMID: 33460504
- PMCID: PMC8359336
- DOI: 10.1111/febs.15715
More than just protein building blocks: how amino acids and related metabolic pathways fuel macrophage polarization
Abstract
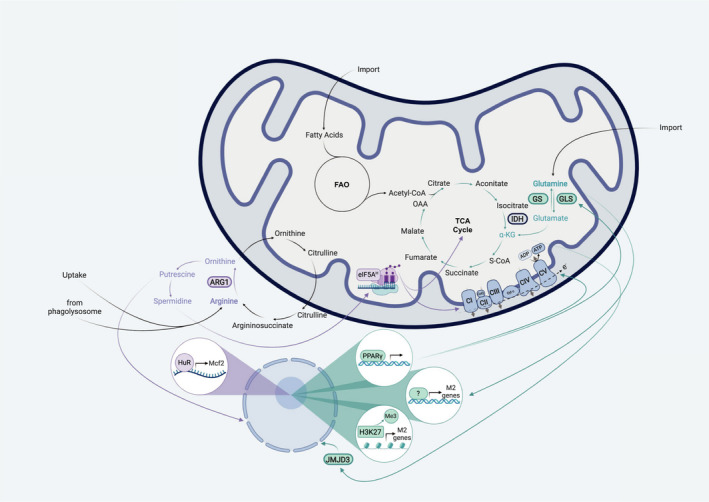
Macrophages represent the first line of defence in innate immune responses and additionally serve important functions for the regulation of host inflammation and tissue homeostasis. The M1/M2 model describes the two extremes of macrophage polarization states, which can be induced by multiple stimuli, most notably by LPS/IFN-γ and IL-4/IL-13. Historically, the expression of two genes encoding for enzymes, which use the same amino acid as their substrate, iNOS and ARG1, has been used to define classically activated M1 (iNOS) and alternatively activated M2 (ARG1) macrophages. This 'arginine dichotomy' has recently become a matter of debate; however, in parallel with the emerging field of immunometabolism there is accumulating evidence that these two enzymes and their related metabolites are fundamentally involved in the intrinsic regulation of macrophage polarization and function. The aim of this review is to highlight recent advances in macrophage biology and immunometabolism with a specific focus on amino acid metabolism and their related metabolic pathways: iNOS/ARG1 (arginine), TCA cycle and OXPHOS (glutamine) as well as the one-carbon metabolism (serine, glycine).
Keywords: TCA cycle; arginase/iNOS; glutamine; immunometabolism; macrophage polarization; nitric oxide; oxidative phosphorylation; polyamines; serine; α-ketoglutarate.
© 2021 The Authors. The FEBS Journal published by John Wiley & Sons Ltd on behalf of Federation of European Biochemical Societies.
Conflict of interest statement
The authors declare no conflict of interest.
Figures

Similar articles
-
Grossamide attenuates inflammation by balancing macrophage polarization through metabolic reprogramming of macrophages in mice.Int Immunopharmacol. 2022 Nov;112:109190. doi: 10.1016/j.intimp.2022.109190. Epub 2022 Sep 15. Int Immunopharmacol. 2022. PMID: 36116152
-
[The modulation of low-level laser on polarization of mouse bone marrow-derived macrophages].Xi Bao Yu Fen Zi Mian Yi Xue Za Zhi. 2016 Aug;32(8):1045-50. Xi Bao Yu Fen Zi Mian Yi Xue Za Zhi. 2016. PMID: 27412934 Chinese.
-
[Adipose-derived stem cells promote the polarization from M1 macrophages to M2 macrophages].Xi Bao Yu Fen Zi Mian Yi Xue Za Zhi. 2016 Mar;32(3):332-8. Xi Bao Yu Fen Zi Mian Yi Xue Za Zhi. 2016. PMID: 26927552 Chinese.
-
Heterogeneity of macrophage activation in fish.Dev Comp Immunol. 2011 Dec;35(12):1246-55. doi: 10.1016/j.dci.2011.03.008. Epub 2011 Mar 23. Dev Comp Immunol. 2011. PMID: 21414343 Review.
-
Macrophage Polarization: Different Gene Signatures in M1(LPS+) vs. Classically and M2(LPS-) vs. Alternatively Activated Macrophages.Front Immunol. 2019 May 24;10:1084. doi: 10.3389/fimmu.2019.01084. eCollection 2019. Front Immunol. 2019. PMID: 31178859 Free PMC article. Review.
Cited by
-
Emerging nanomaterials targeting macrophage adapted to abnormal metabolism in cancer and atherosclerosis therapy (Review).Int J Mol Med. 2024 Feb;53(2):13. doi: 10.3892/ijmm.2023.5337. Epub 2023 Dec 8. Int J Mol Med. 2024. PMID: 38063240 Free PMC article. Review.
-
The levels of amino acid metabolites in serum induce the pathogenesis of atopic dermatitis by mediating the inflammatory protein S100A12.Sci Rep. 2024 Oct 8;14(1):23435. doi: 10.1038/s41598-024-74522-1. Sci Rep. 2024. PMID: 39379513 Free PMC article.
-
Primary Human M2 Macrophage Subtypes Are Distinguishable by Aqueous Metabolite Profiles.Int J Mol Sci. 2024 Feb 18;25(4):2407. doi: 10.3390/ijms25042407. Int J Mol Sci. 2024. PMID: 38397084 Free PMC article.
-
Roles of Critical Amino Acids Metabolism in The Interactions Between Intracellular Bacterial Infection and Macrophage Function.Curr Microbiol. 2024 Jul 20;81(9):280. doi: 10.1007/s00284-024-03801-x. Curr Microbiol. 2024. PMID: 39031203 Review.
-
Macrophage polarisation and inflammatory mechanisms in atherosclerosis: Implications for prevention and treatment.Heliyon. 2024 May 29;10(11):e32073. doi: 10.1016/j.heliyon.2024.e32073. eCollection 2024 Jun 15. Heliyon. 2024. PMID: 38873669 Free PMC article. Review.
References
-
- Warburg O, Gawehn K & Geissler AW (1958) Metabolism of leukocytes. Zeitschrift fur Naturforschung Teil B, Chemie, Biochemie, Biophysik, Biologie und verwandte Gebiete 13b, 515–516. - PubMed
-
- Levene P & Meyer G (1912) On the action of leucocytes on glucose second communication. J Biol Chem 12, 265–273.
-
- Levene PA & Meyer GM (1912) THE action of leucocytes on glucose. J Biol Chem 11, 361–370.
-
- Fleischmann W & Kubowitz F (1927) Über den stoffwechsel der leukocyten. Biochem Z 181, 395.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
