

Adipose tissue inflammation and systemic insulin resistance in mice with diet-induced obesity is possibly associated with disruption of PFKFB3 in hematopoietic cells
- PMID: 33462362
- PMCID: PMC7897240
- DOI: 10.1038/s41374-020-00523-z
Adipose tissue inflammation and systemic insulin resistance in mice with diet-induced obesity is possibly associated with disruption of PFKFB3 in hematopoietic cells
Abstract
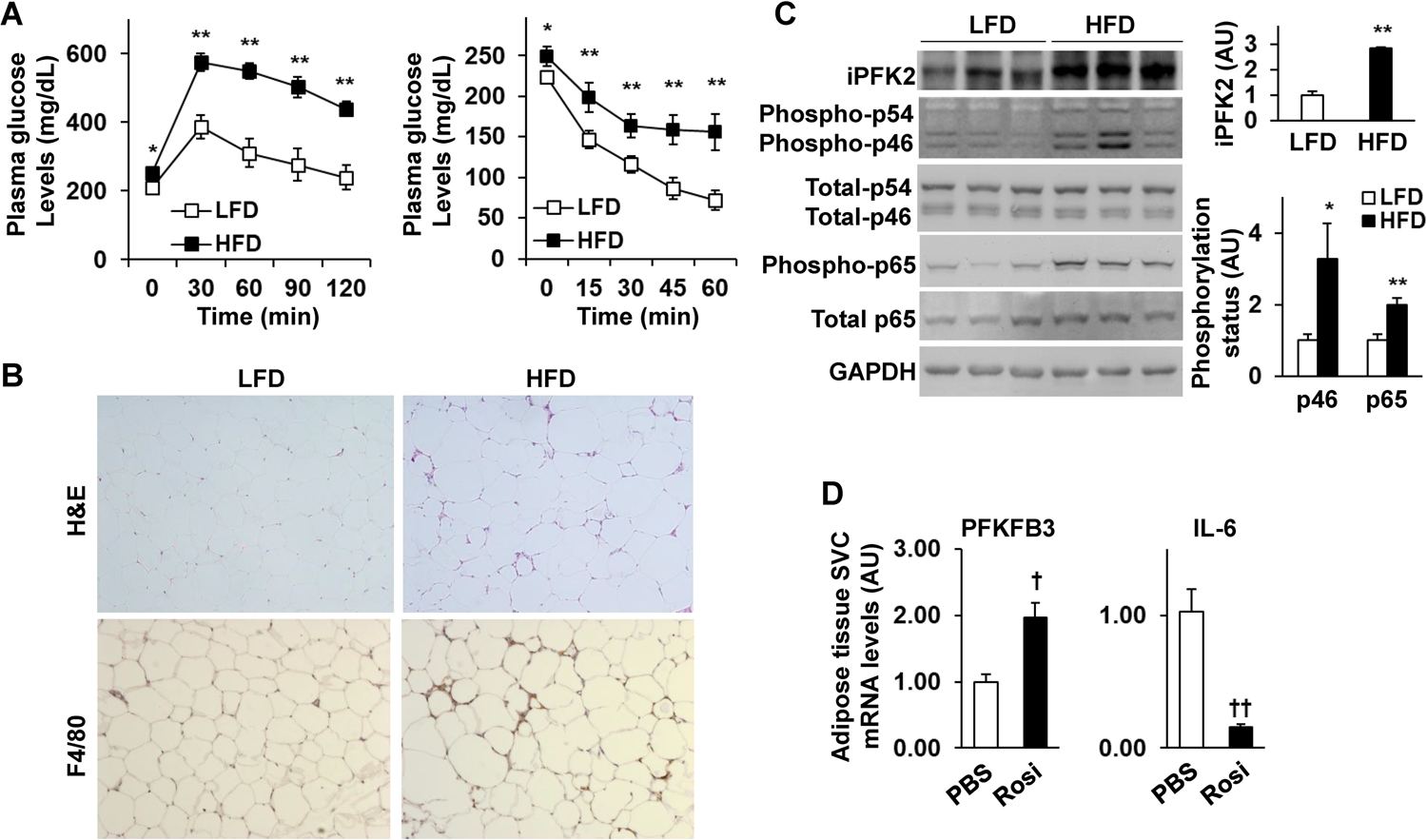
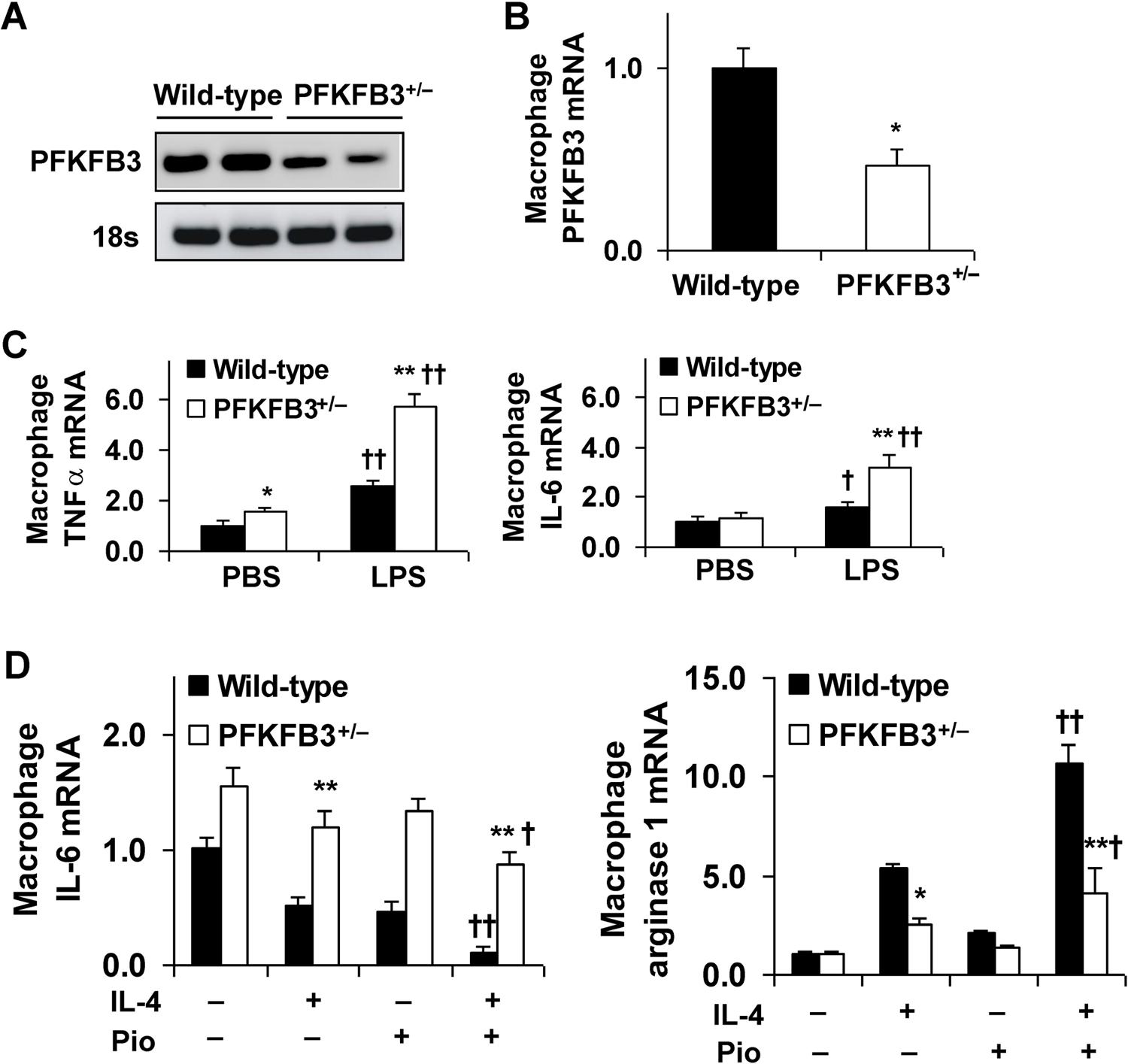
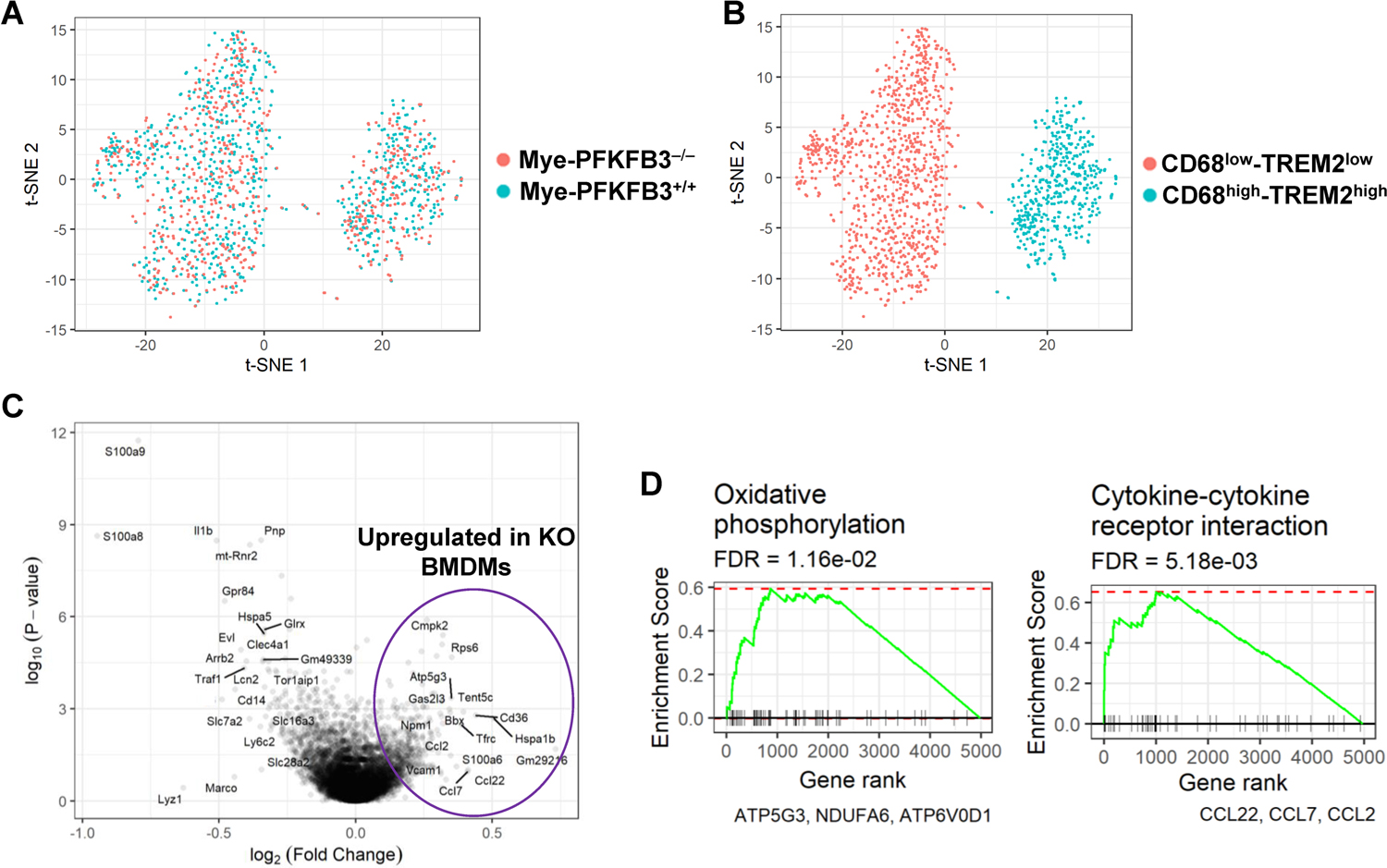
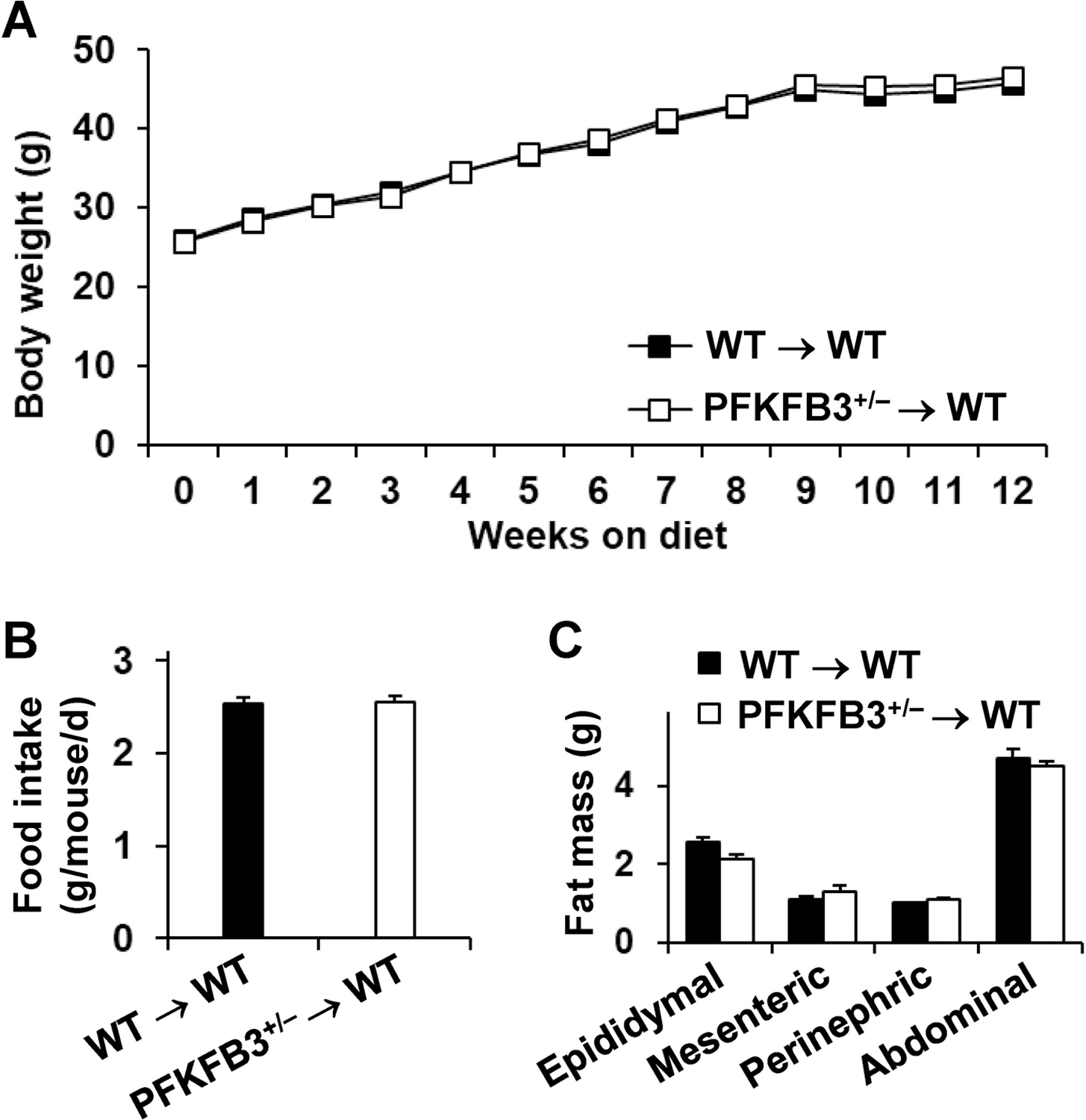
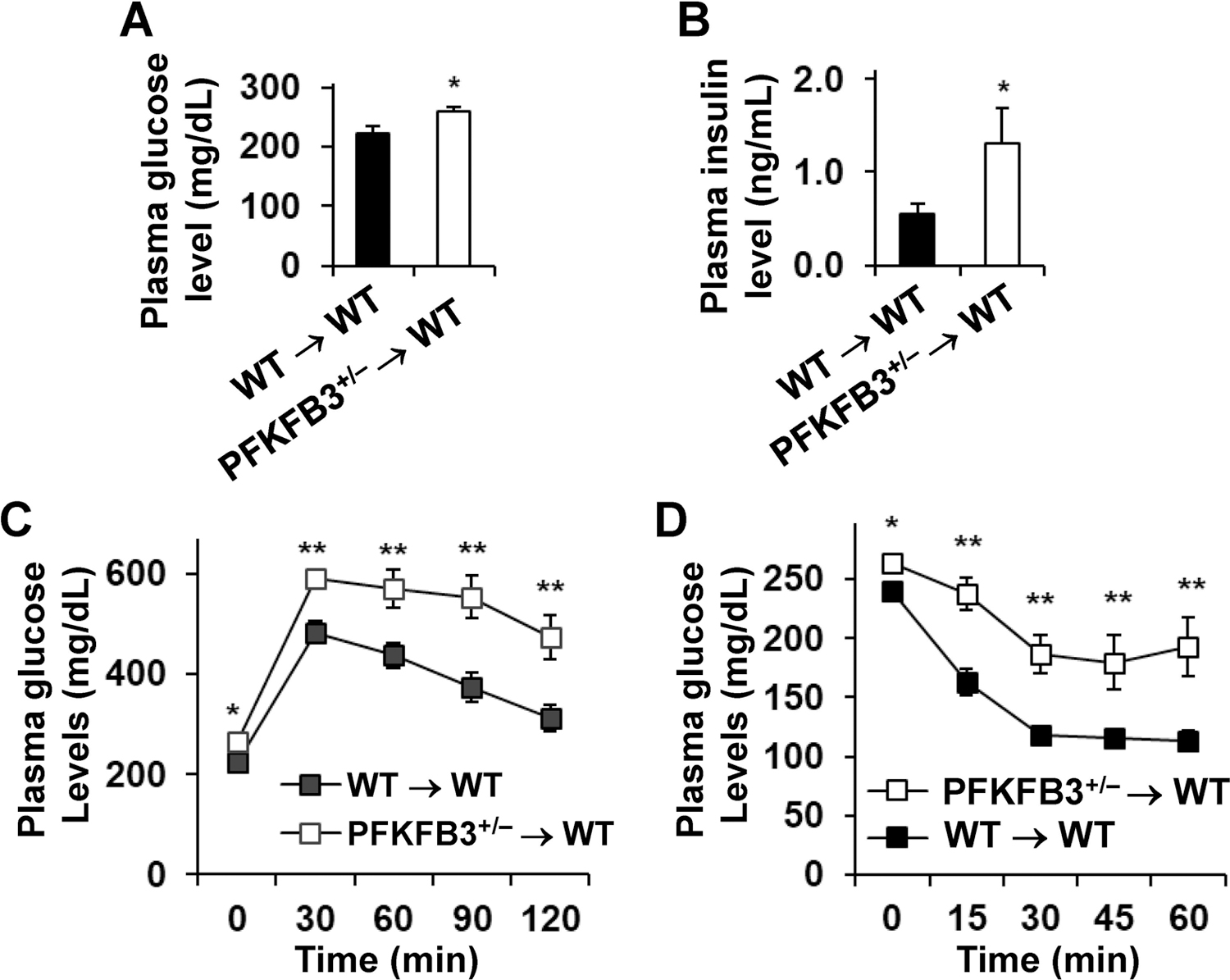
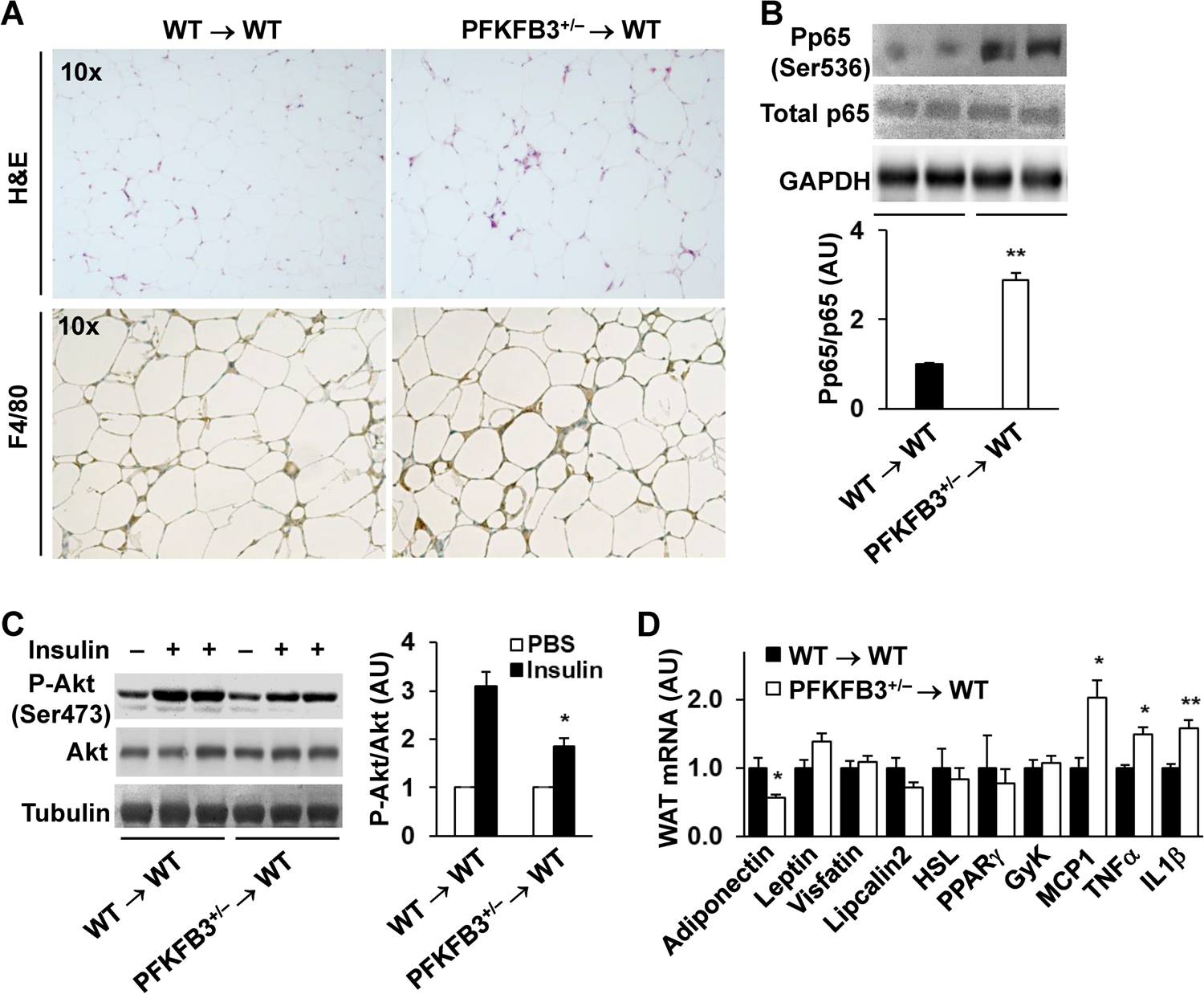
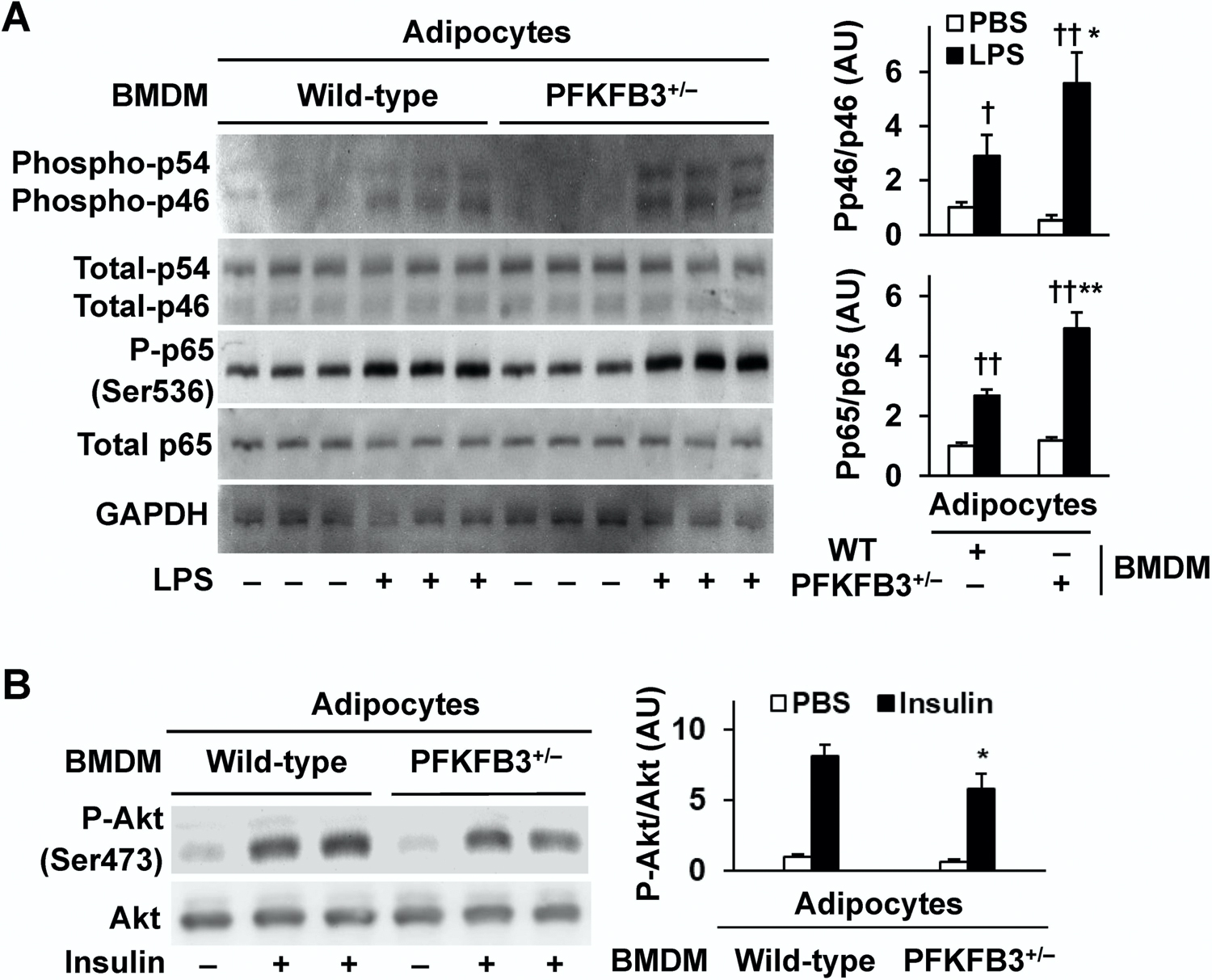
Obesity-associated inflammation in white adipose tissue (WAT) is a causal factor of systemic insulin resistance; however, precisely how immune cells regulate WAT inflammation in relation to systemic insulin resistance remains to be elucidated. The present study examined a role for 6-phosphofructo-2-kinase/fructose-2,6-bisphosphatase 3 (PFKFB3) in hematopoietic cells in regulating WAT inflammation and systemic insulin sensitivity. Male C57BL/6J mice were fed a high-fat diet (HFD) or low-fat diet (LFD) for 12 weeks and examined for WAT inducible 6-phosphofructo-2-kinase (iPFK2) content, while additional HFD-fed mice were treated with rosiglitazone and examined for PFKFB3 mRNAs in WAT stromal vascular cells (SVC). Also, chimeric mice in which PFKFB3 was disrupted only in hematopoietic cells and control chimeric mice were also fed an HFD and examined for HFD-induced WAT inflammation and systemic insulin resistance. In vitro, adipocytes were co-cultured with bone marrow-derived macrophages and examined for adipocyte proinflammatory responses and insulin signaling. Compared with their respective levels in controls, WAT iPFK2 amount in HFD-fed mice and WAT SVC PFKFB3 mRNAs in rosiglitazone-treated mice were significantly increased. When the inflammatory responses were analyzed, peritoneal macrophages from PFKFB3-disrputed mice revealed increased proinflammatory activation and decreased anti-inflammatory activation compared with control macrophages. At the whole animal level, hematopoietic cell-specific PFKFB3 disruption enhanced the effects of HFD feeding on promoting WAT inflammation, impairing WAT insulin signaling, and increasing systemic insulin resistance. In vitro, adipocytes co-cultured with PFKFB3-disrupted macrophages revealed increased proinflammatory responses and decreased insulin signaling compared with adipocytes co-cultured with control macrophages. These results suggest that PFKFB3 disruption in hematopoietic cells only exacerbates HFD-induced WAT inflammation and systemic insulin resistance.
Conflict of interest statement
The authors declare that they do not have conflict of interest.
Conflict of Interest
The authors do not have conflict of interest.
Figures
Similar articles
-
Adipocyte inducible 6-phosphofructo-2-kinase suppresses adipose tissue inflammation and promotes macrophage anti-inflammatory activation.J Nutr Biochem. 2021 Sep;95:108764. doi: 10.1016/j.jnutbio.2021.108764. Epub 2021 May 6. J Nutr Biochem. 2021. PMID: 33964465 Free PMC article.
-
Disruption of inducible 6-phosphofructo-2-kinase impairs the suppressive effect of PPARγ activation on diet-induced intestine inflammatory response.J Nutr Biochem. 2013 May;24(5):770-5. doi: 10.1016/j.jnutbio.2012.04.007. Epub 2012 Jul 26. J Nutr Biochem. 2013. PMID: 22841546 Free PMC article.
-
Disruption of inducible 6-phosphofructo-2-kinase ameliorates diet-induced adiposity but exacerbates systemic insulin resistance and adipose tissue inflammatory response.J Biol Chem. 2010 Feb 5;285(6):3713-3721. doi: 10.1074/jbc.M109.058446. Epub 2009 Nov 30. J Biol Chem. 2010. PMID: 19948719 Free PMC article.
-
Omega-3 fatty acids and adipose tissue biology.Mol Aspects Med. 2018 Dec;64:147-160. doi: 10.1016/j.mam.2018.01.004. Epub 2018 Jan 17. Mol Aspects Med. 2018. PMID: 29329795 Review.
-
Yin and Yang of hypothalamic insulin and leptin signaling in regulating white adipose tissue metabolism.Rev Endocr Metab Disord. 2011 Sep;12(3):235-43. doi: 10.1007/s11154-011-9190-4. Rev Endocr Metab Disord. 2011. PMID: 21713385 Free PMC article. Review.
Cited by
-
Inhibition of PFKFB Preserves Intestinal Barrier Function in Sepsis by Inhibiting NLRP3/GSDMD.Oxid Med Cell Longev. 2022 Dec 23;2022:8704016. doi: 10.1155/2022/8704016. eCollection 2022. Oxid Med Cell Longev. 2022. PMID: 36589684 Free PMC article.
-
Theabrownin ameliorates liver inflammation, oxidative stress, and fibrosis in MCD diet-fed C57BL/6J mice.Front Endocrinol (Lausanne). 2023 Jan 18;14:1118925. doi: 10.3389/fendo.2023.1118925. eCollection 2023. Front Endocrinol (Lausanne). 2023. PMID: 36742397 Free PMC article.
-
Molecular and pathophysiological relationship between obesity and chronic inflammation in the manifestation of metabolic dysfunctions and their inflammation‑mediating treatment options (Review).Mol Med Rep. 2024 Jun;29(6):95. doi: 10.3892/mmr.2024.13219. Epub 2024 Apr 12. Mol Med Rep. 2024. PMID: 38606791 Free PMC article. Review.
-
Different Effects of Leucine Supplementation and/or Exercise on Systemic Insulin Sensitivity in Mice.Front Endocrinol (Lausanne). 2021 May 12;12:651303. doi: 10.3389/fendo.2021.651303. eCollection 2021. Front Endocrinol (Lausanne). 2021. PMID: 34054726 Free PMC article.
-
Role of PFKFB3-driven glycolysis in sepsis.Ann Med. 2023 Dec;55(1):1278-1289. doi: 10.1080/07853890.2023.2191217. Ann Med. 2023. PMID: 37199341 Free PMC article. Review.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
