

Characterizing the portability of phage-encoded homologous recombination proteins
- PMID: 33462496
- PMCID: PMC7990699
- DOI: 10.1038/s41589-020-00710-5
Characterizing the portability of phage-encoded homologous recombination proteins
Abstract
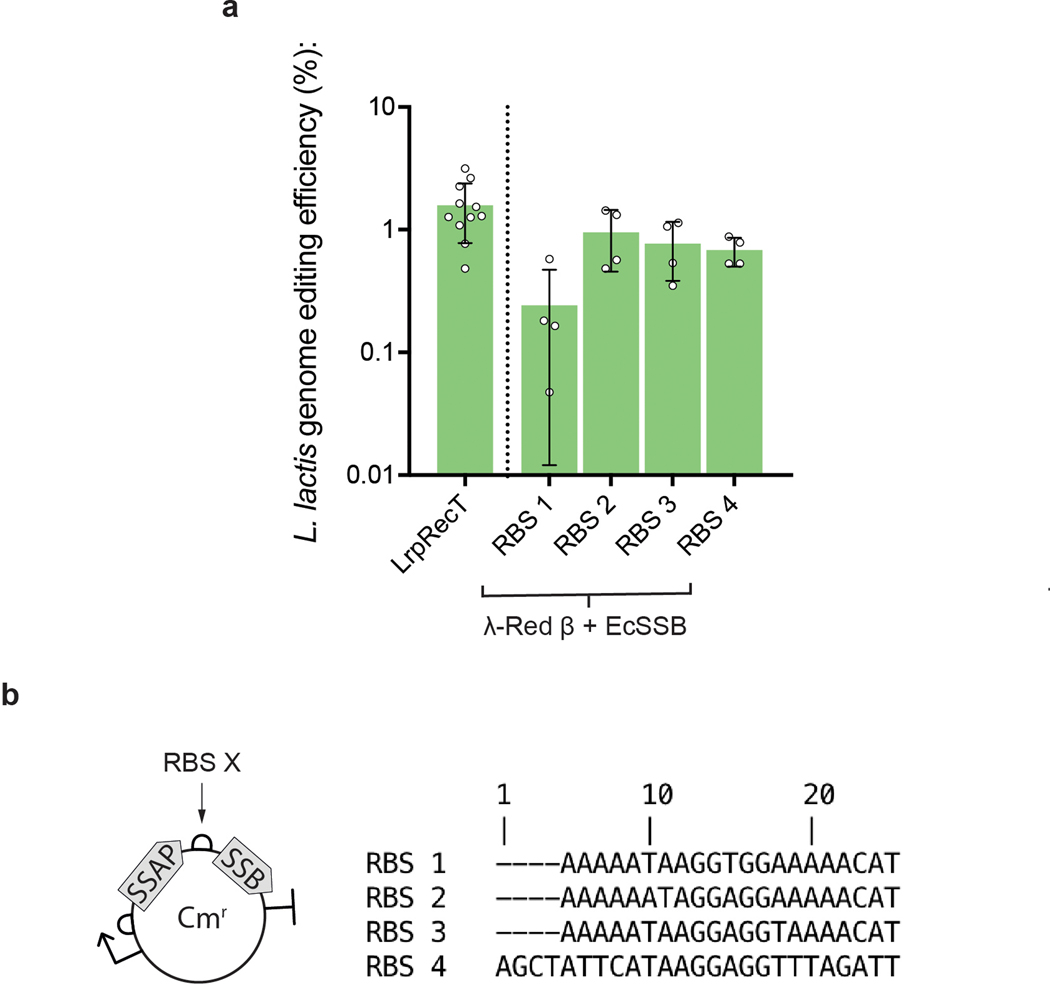
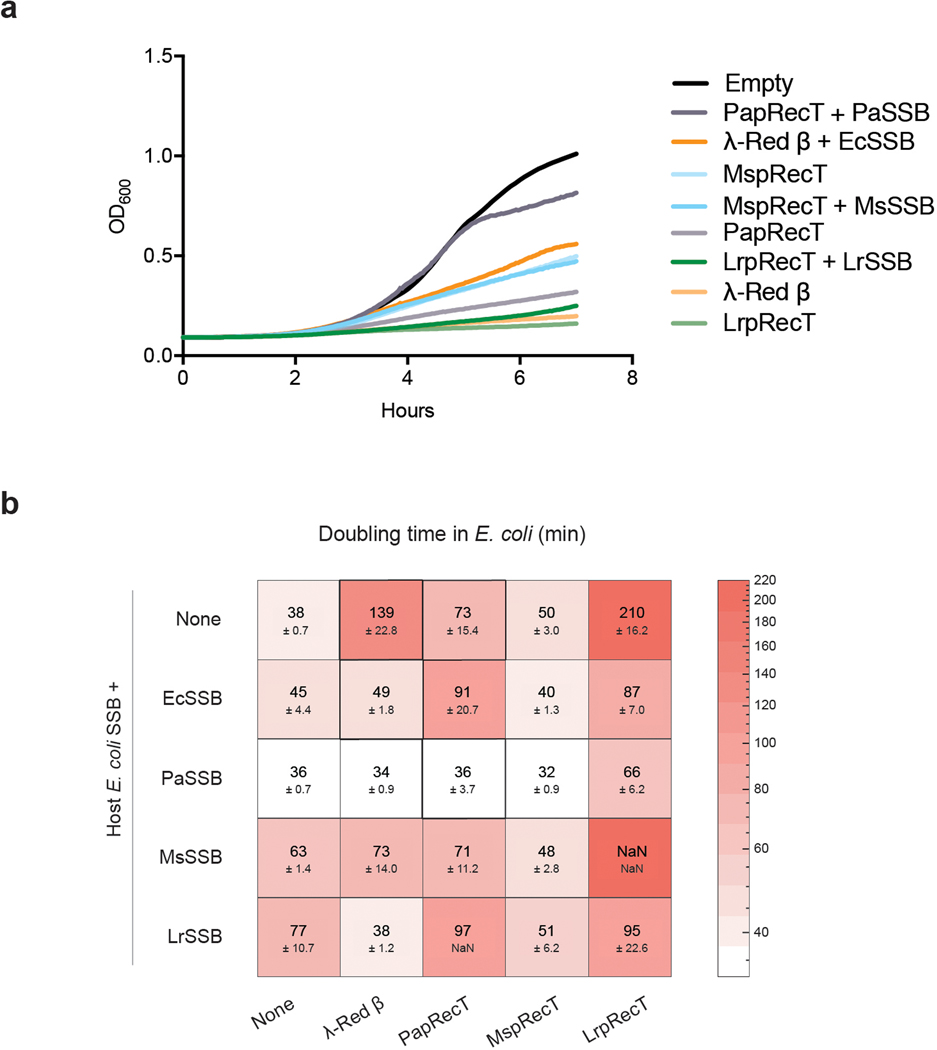
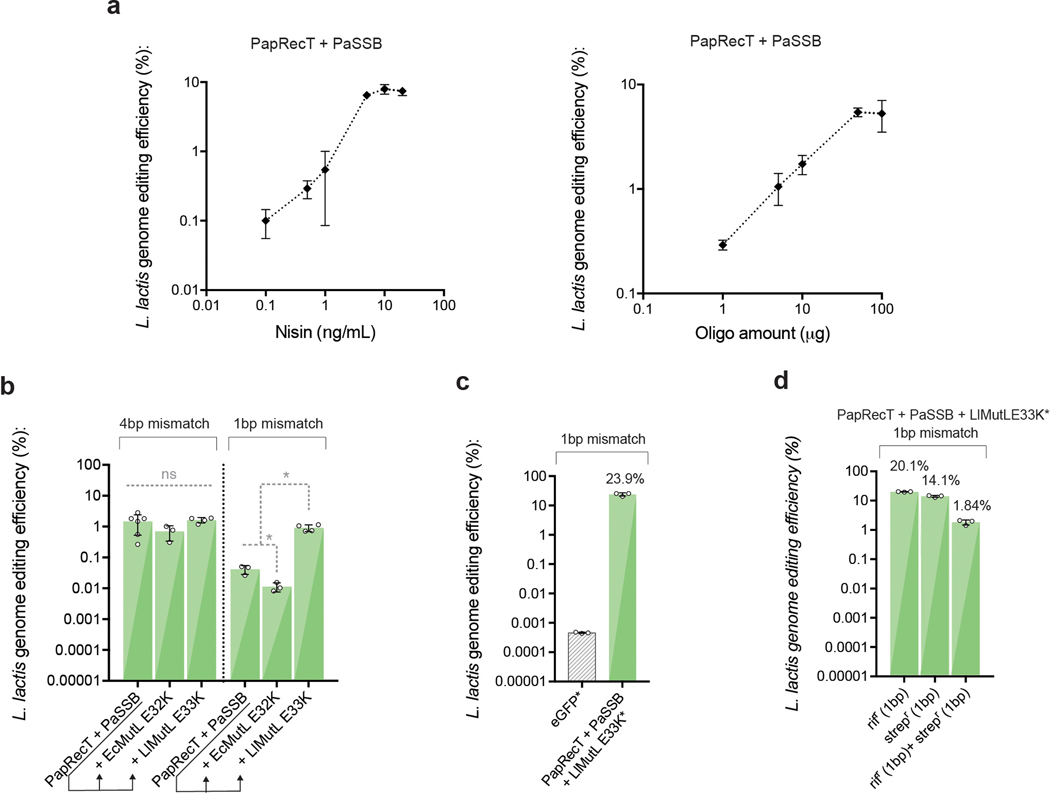
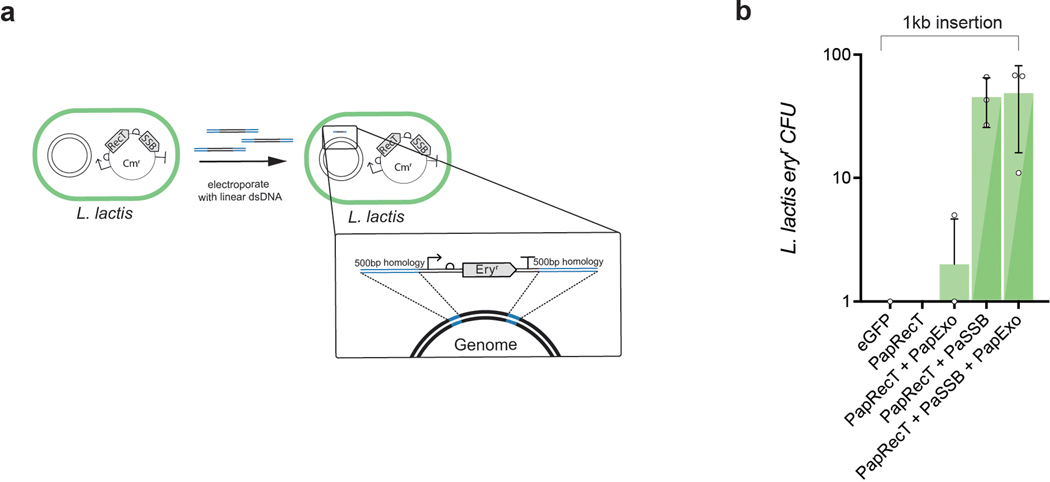
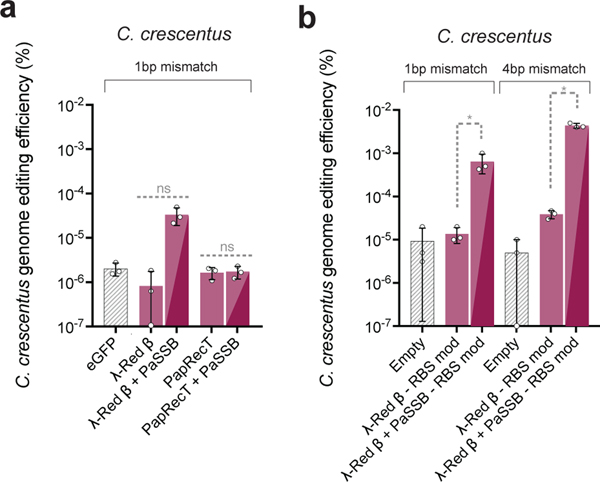
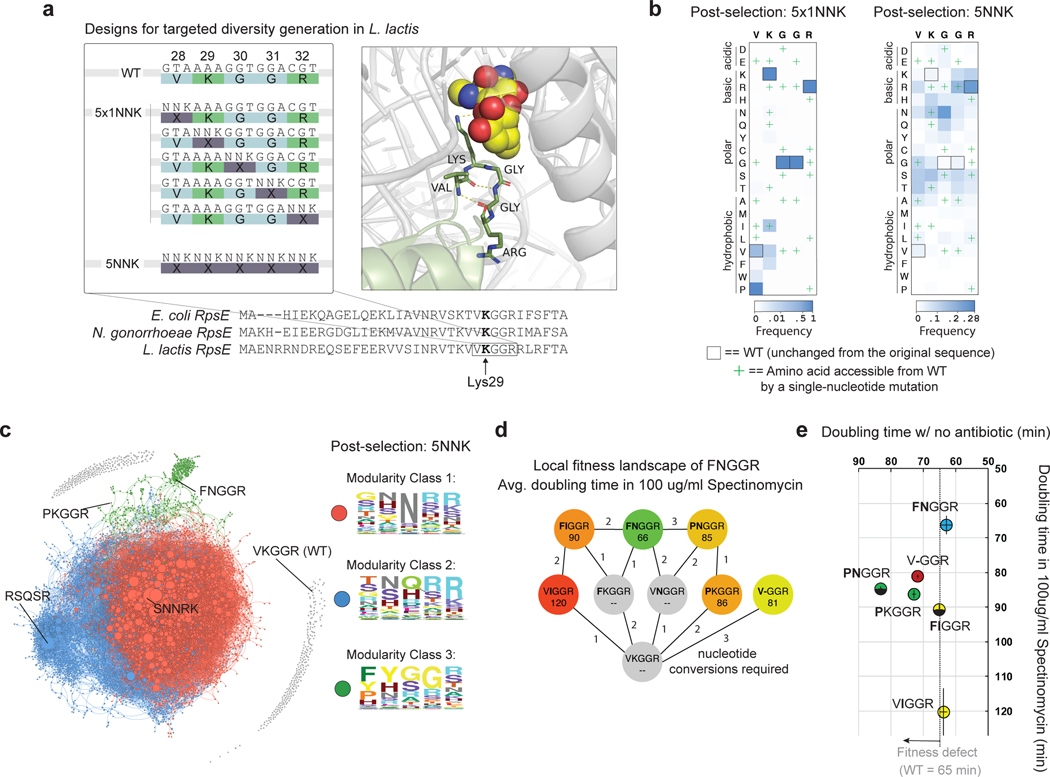
Efficient genome editing methods are essential for biotechnology and fundamental research. Homologous recombination (HR) is the most versatile method of genome editing, but techniques that rely on host RecA-mediated pathways are inefficient and laborious. Phage-encoded single-stranded DNA annealing proteins (SSAPs) improve HR 1,000-fold above endogenous levels. However, they are not broadly functional. Using Escherichia coli, Lactococcus lactis, Mycobacterium smegmatis, Lactobacillus rhamnosus and Caulobacter crescentus, we investigated the limited portability of SSAPs. We find that these proteins specifically recognize the C-terminal tail of the host's single-stranded DNA-binding protein (SSB) and are portable between species only if compatibility with this host domain is maintained. Furthermore, we find that co-expressing SSAPs with SSBs can significantly improve genome editing efficiency, in some species enabling SSAP functionality even without host compatibility. Finally, we find that high-efficiency HR far surpasses the mutational capacity of commonly used random mutagenesis methods, generating exceptional phenotypes that are inaccessible through sequential nucleotide conversions.
Conflict of interest statement
Figures
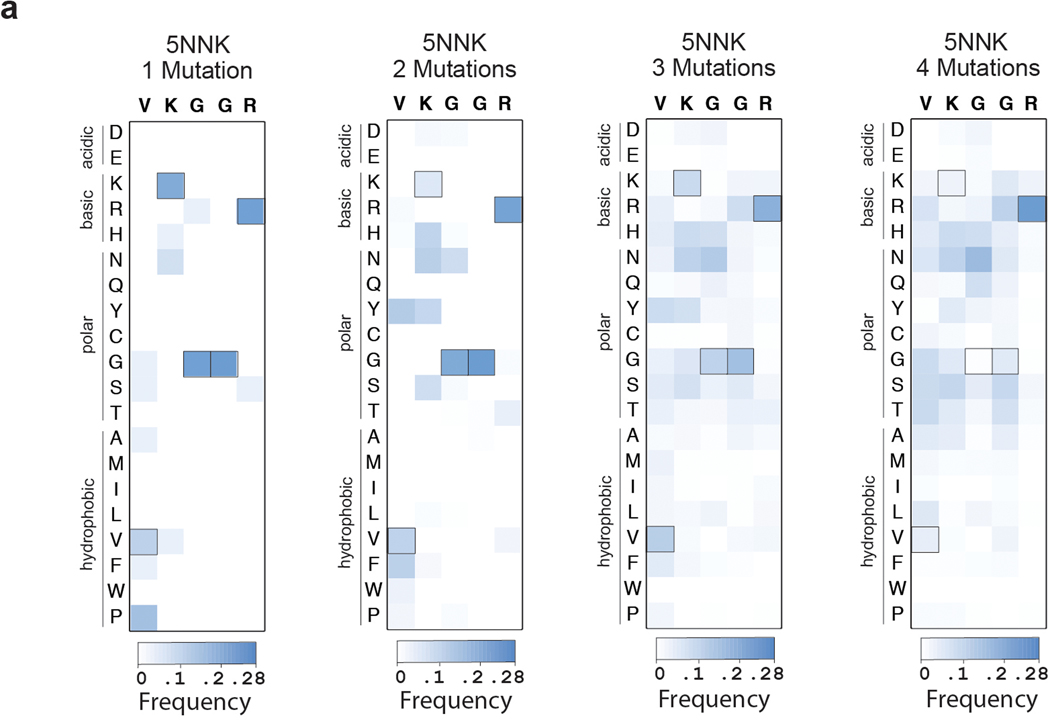
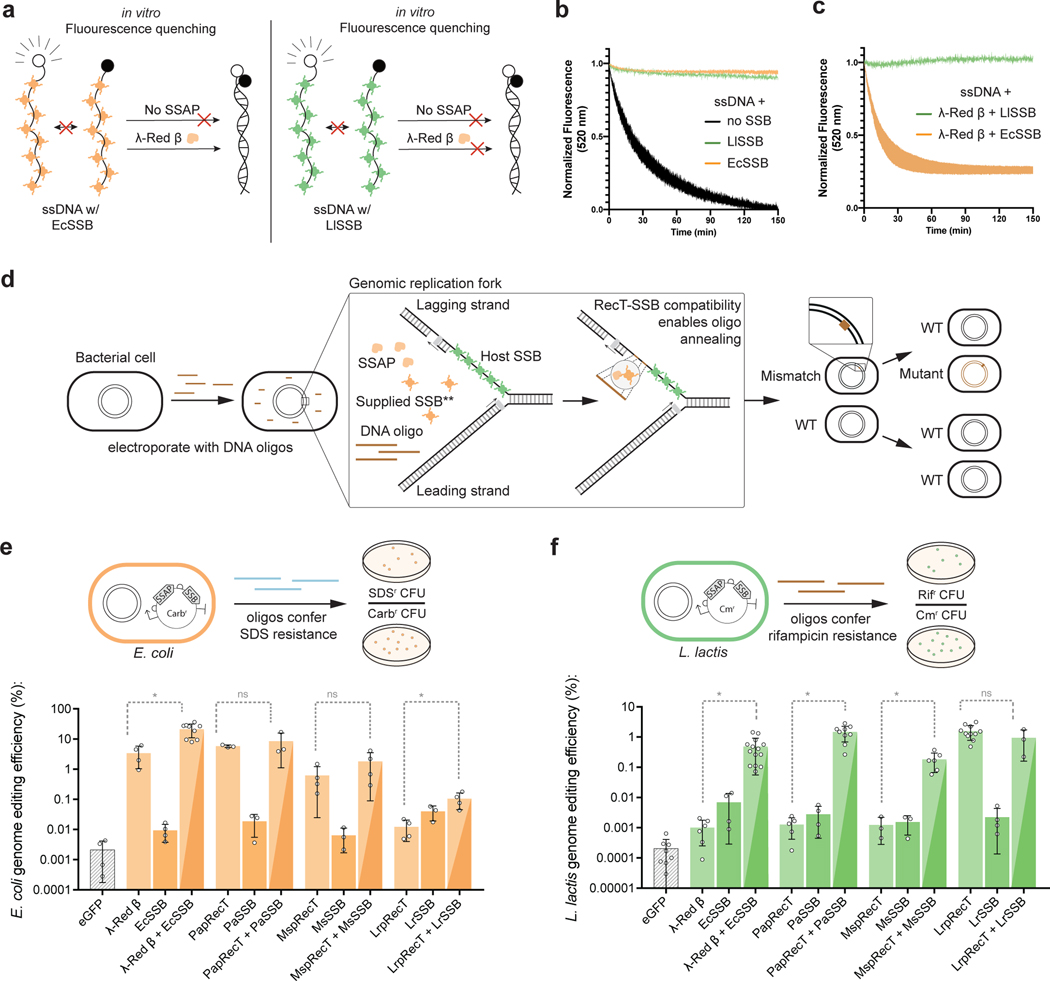
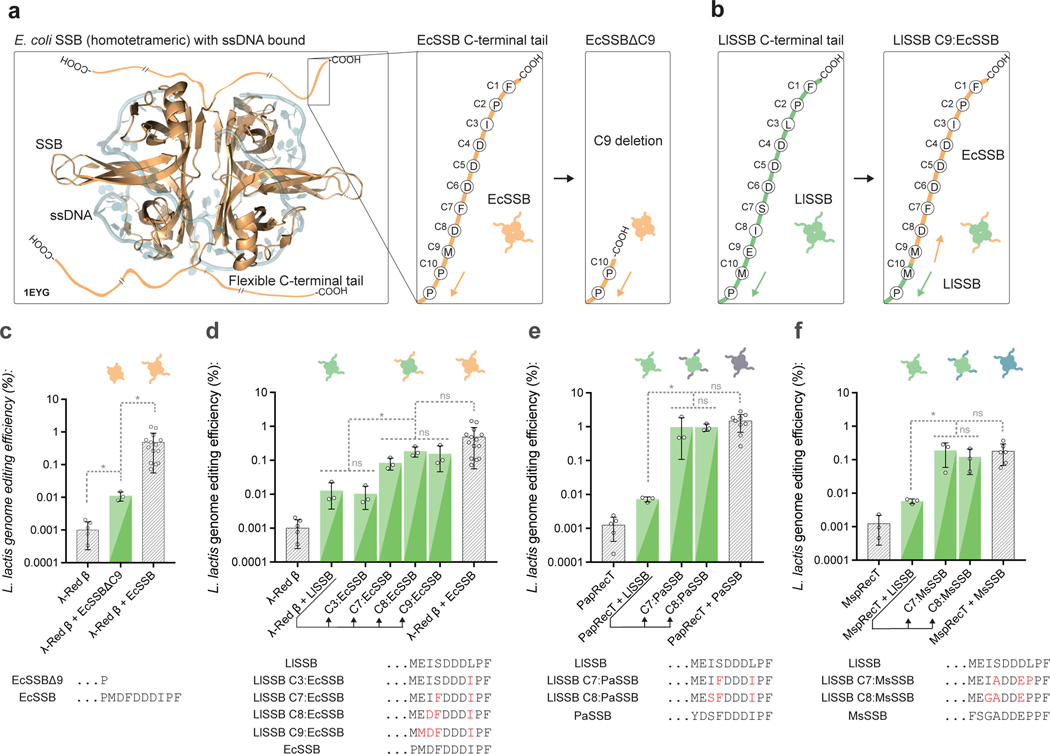
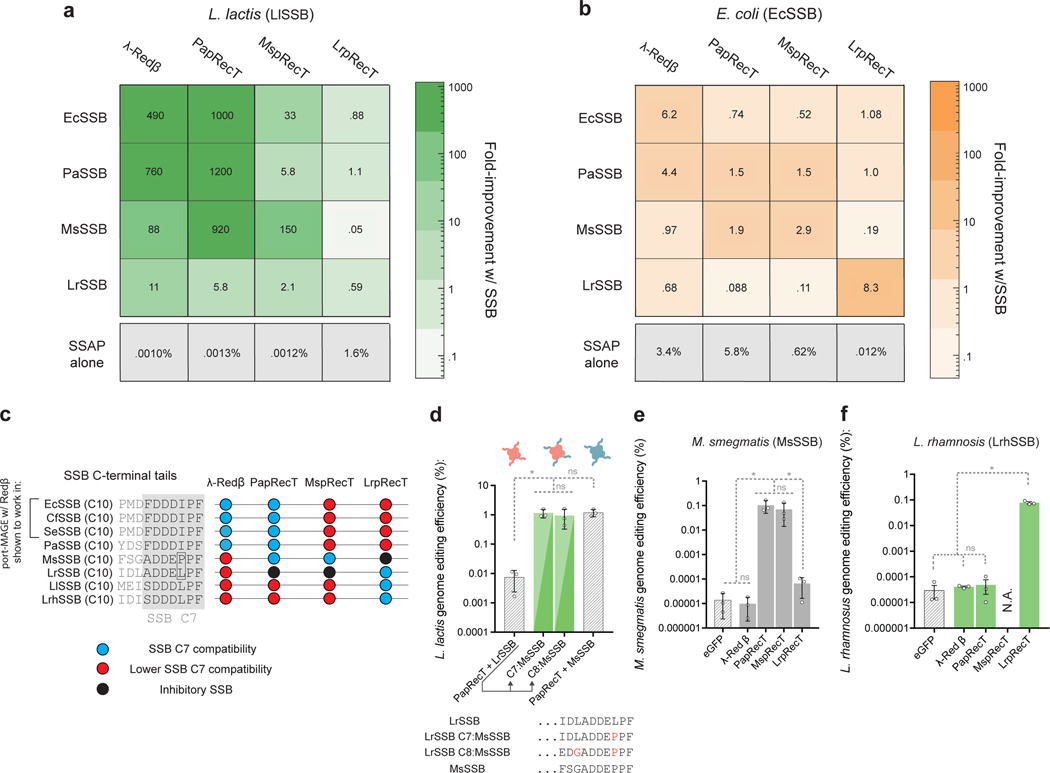
References
-
- Steidler L et al. Biological containment of genetically modified Lactococcus lactis for intestinal delivery of human interleukin 10. Nature Biotechnology 21, 785–789 (2003). - PubMed
Methods-only References:
-
- Choe W, Chandrasegaran S & Ostermeier M Protein fragment complementation in M.HhaI DNA methyltransferase. Biochemical and Biophysical Research Communications 334, 1233–1240 (2005). - PubMed
-
- Lohman TM, Green JM & Beyer RS Large-scale overproduction and rapid purification of the Escherichia coli ssb gene product. Expression of the ssb gene under .lambda. PL control. Biochemistry 25, 21–25 (1986). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
