

Mapping and role of T cell response in SARS-CoV-2-infected mice
- PMID: 33464307
- PMCID: PMC7814348
- DOI: 10.1084/jem.20202187
Mapping and role of T cell response in SARS-CoV-2-infected mice
Erratum in
-
Correction: Mapping and role of T cell response in SARS-CoV-2-infected mice.J Exp Med. 2021 Nov 1;218(11):e2020218710052021c. doi: 10.1084/jem.2020218710052021c. Epub 2021 Oct 15. J Exp Med. 2021. PMID: 34653240 Free PMC article. No abstract available.
Abstract
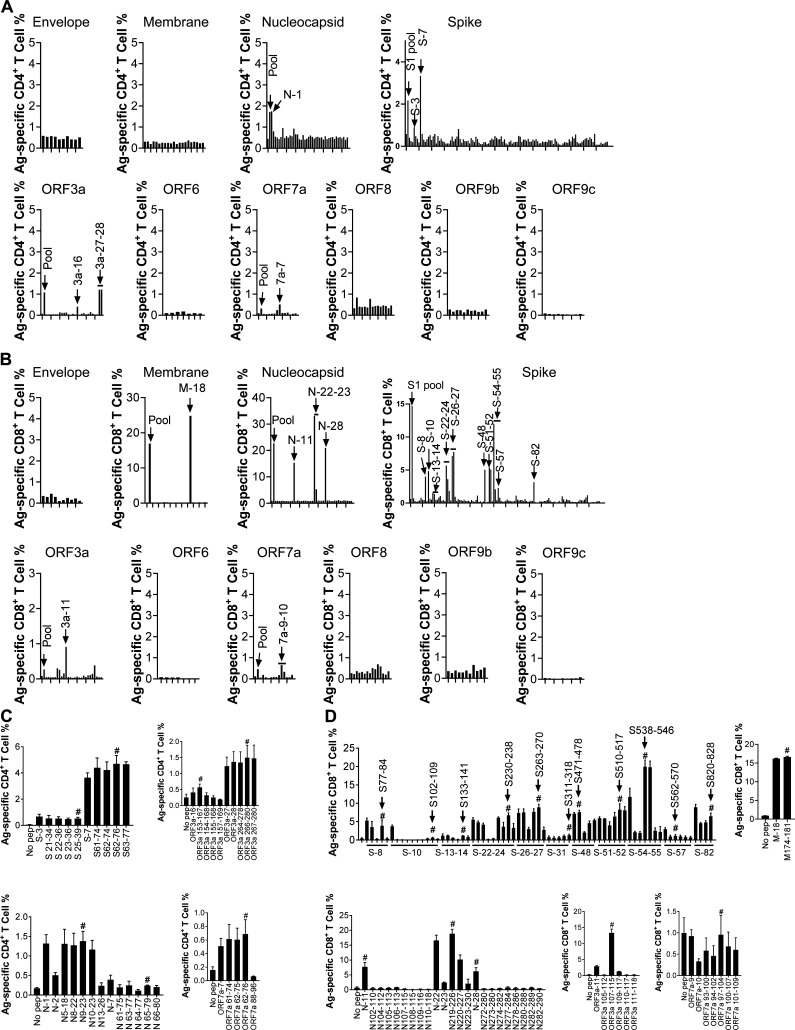
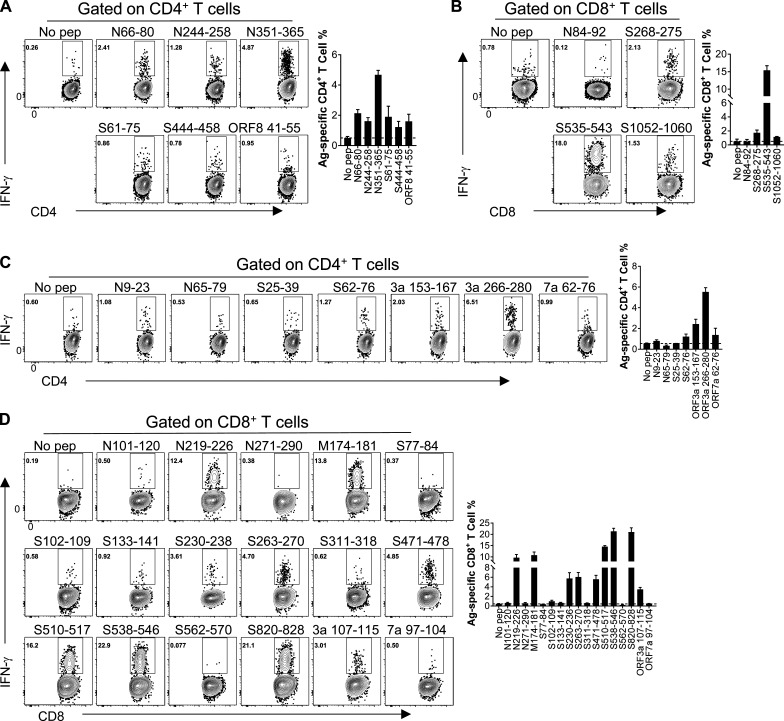
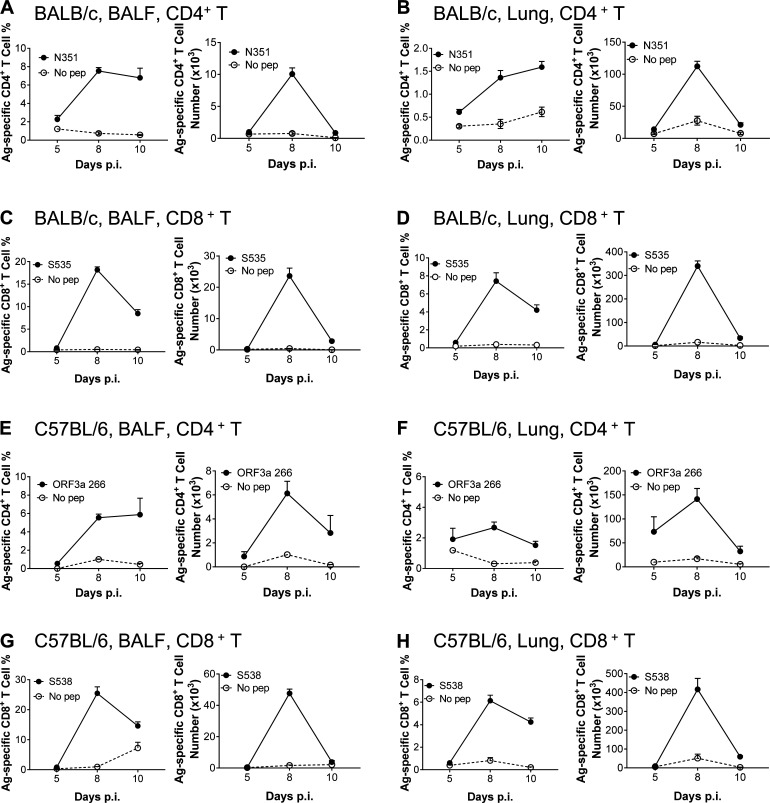
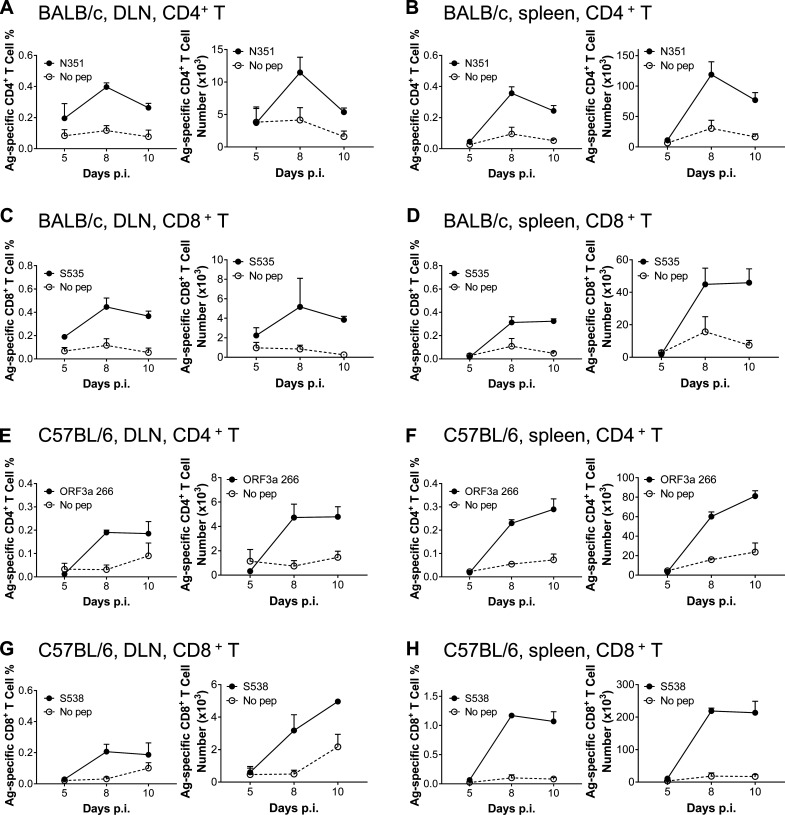
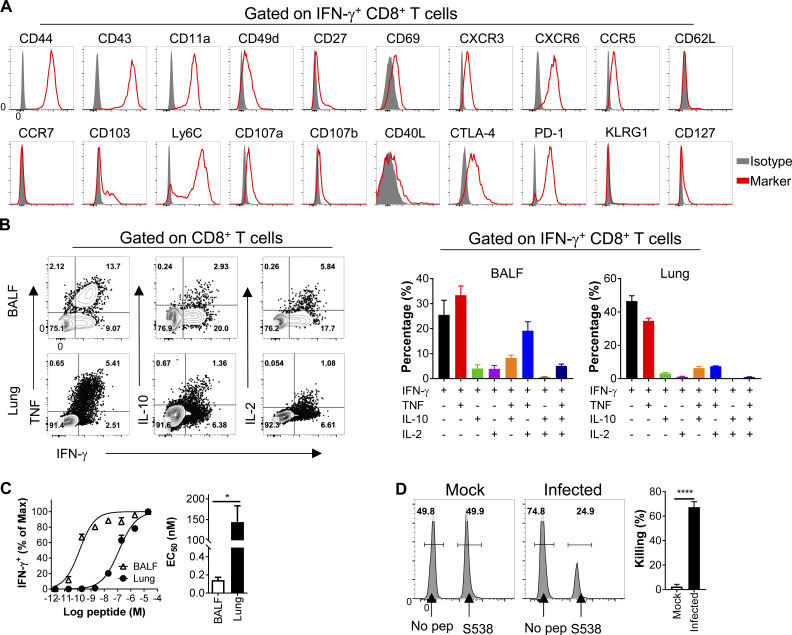
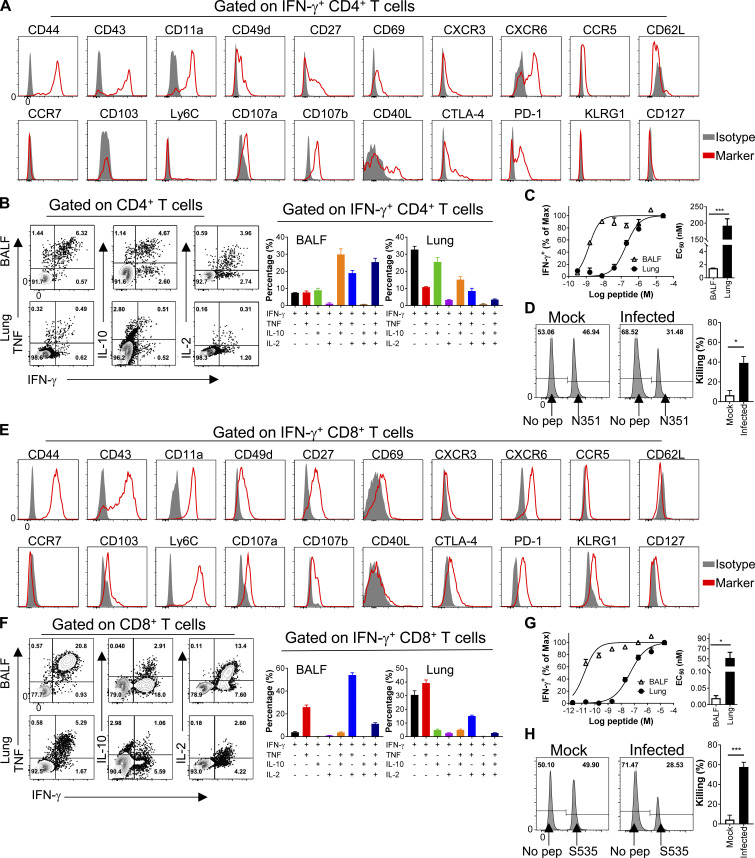
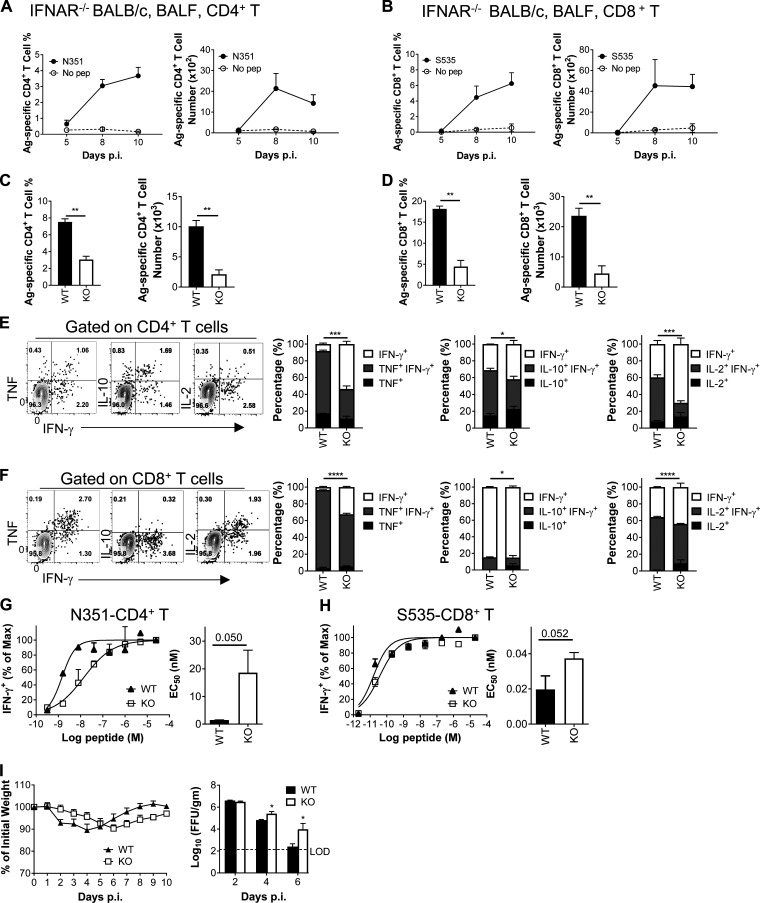
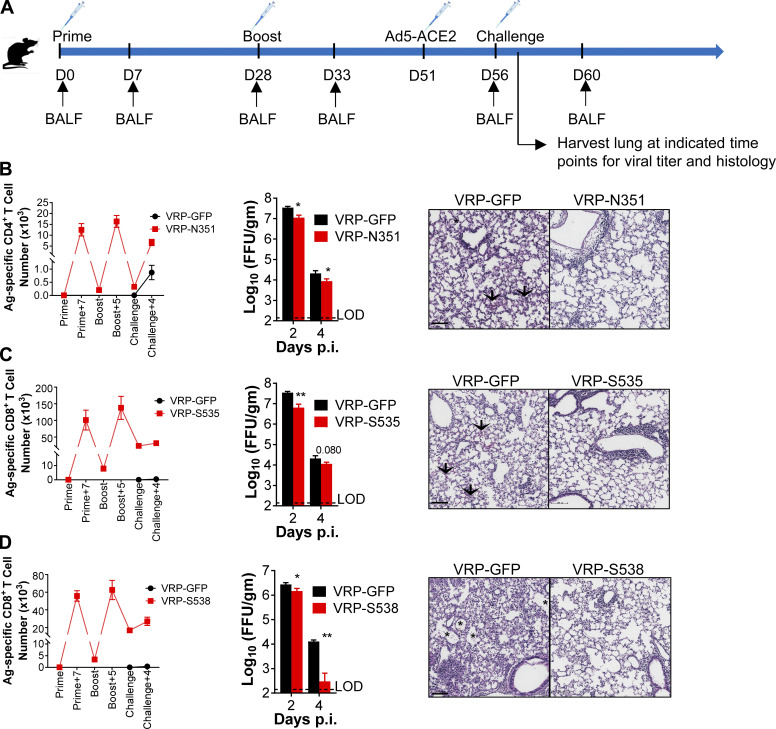
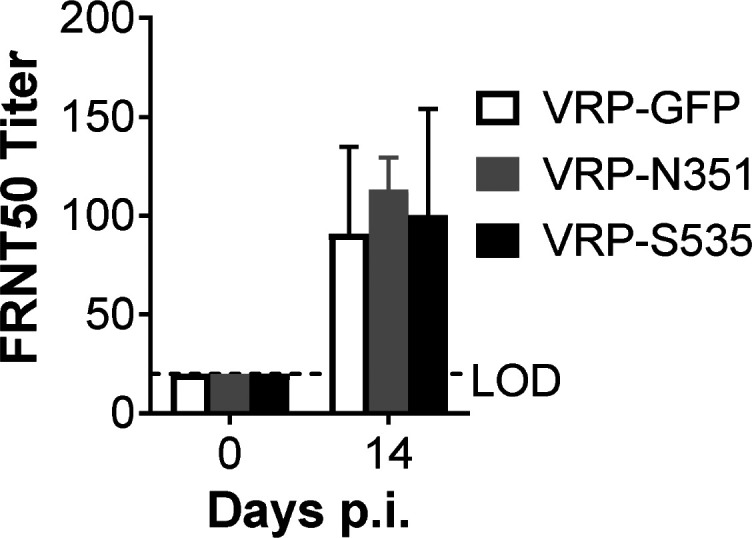
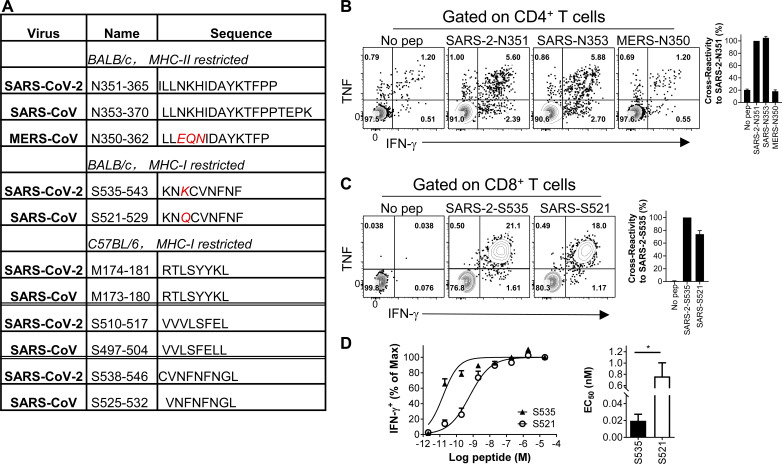
Virus-specific T cells play essential roles in protection against multiple virus infections, including SARS-CoV and MERS-CoV. While SARS-CoV-2-specific T cells have been identified in COVID-19 patients, their role in the protection of SARS-CoV-2-infected mice is not established. Here, using mice sensitized for infection with SARS-CoV-2 by transduction with an adenovirus expressing the human receptor (Ad5-hACE2), we identified SARS-CoV-2-specific T cell epitopes recognized by CD4+ and CD8+ T cells in BALB/c and C57BL/6 mice. Virus-specific T cells were polyfunctional and were able to lyse target cells in vivo. Further, type I interferon pathway was proved to be critical for generating optimal antiviral T cell responses after SARS-CoV-2 infection. T cell vaccination alone partially protected SARS-CoV-2-infected mice from severe disease. In addition, the results demonstrated cross-reactive T cell responses between SARS-CoV and SARS-CoV-2, but not MERS-CoV, in mice. Understanding the role of the T cell response will guide immunopathogenesis studies of COVID-19 and vaccine design and validation.
© 2021 Zhuang et al.
Conflict of interest statement
Disclosures: The authors declare no competing interests exist.
Figures

References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
