

A novel terpene synthase controls differences in anti-aphrodisiac pheromone production between closely related Heliconius butterflies
- PMID: 33465061
- PMCID: PMC7815096
- DOI: 10.1371/journal.pbio.3001022
A novel terpene synthase controls differences in anti-aphrodisiac pheromone production between closely related Heliconius butterflies
Abstract
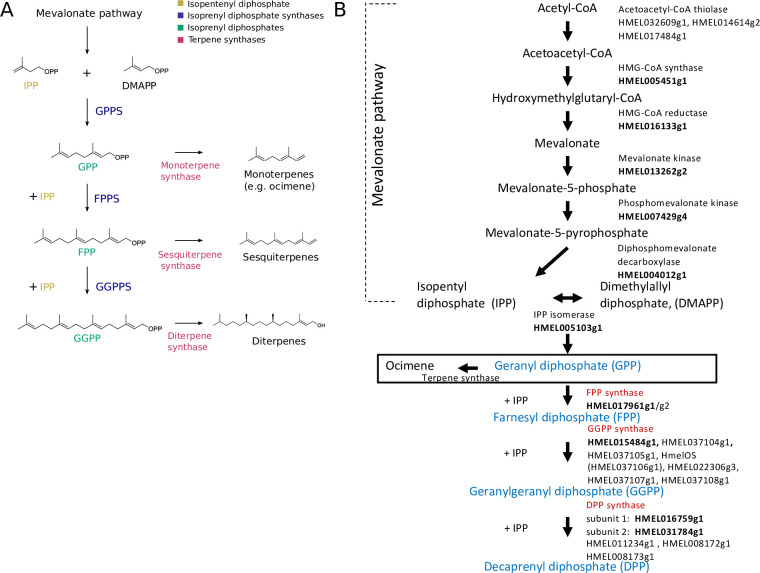
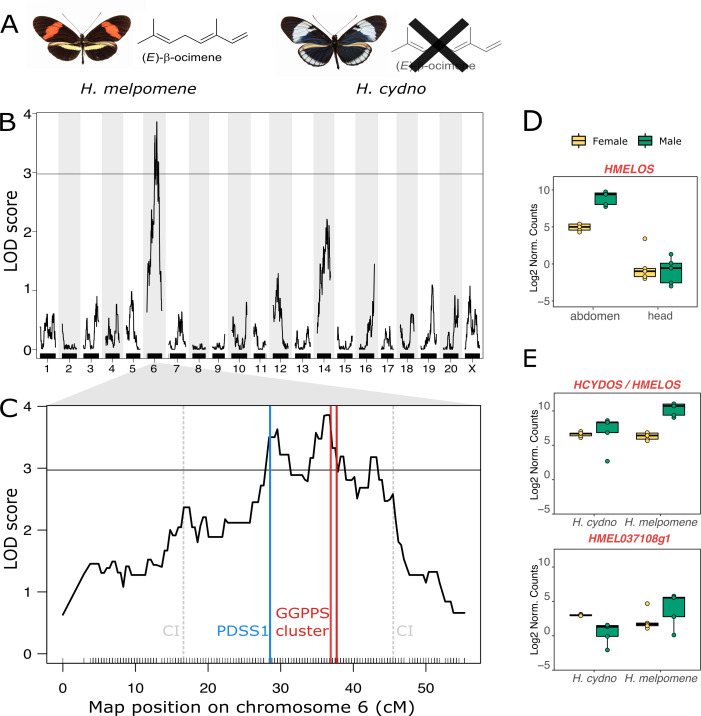
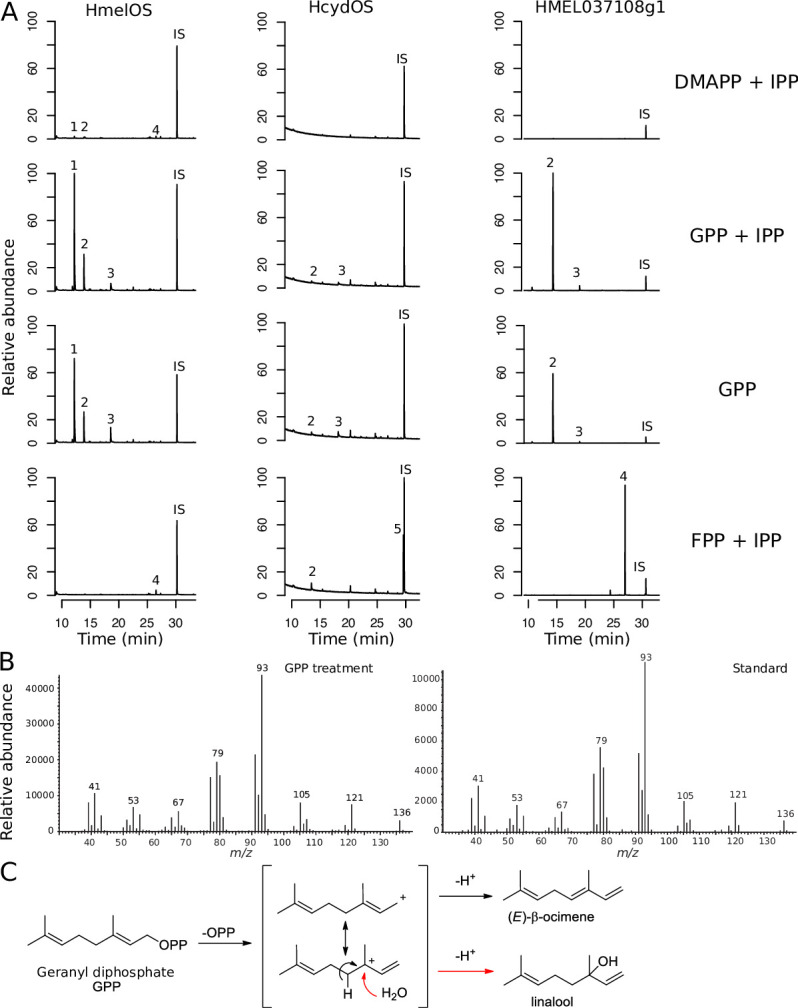
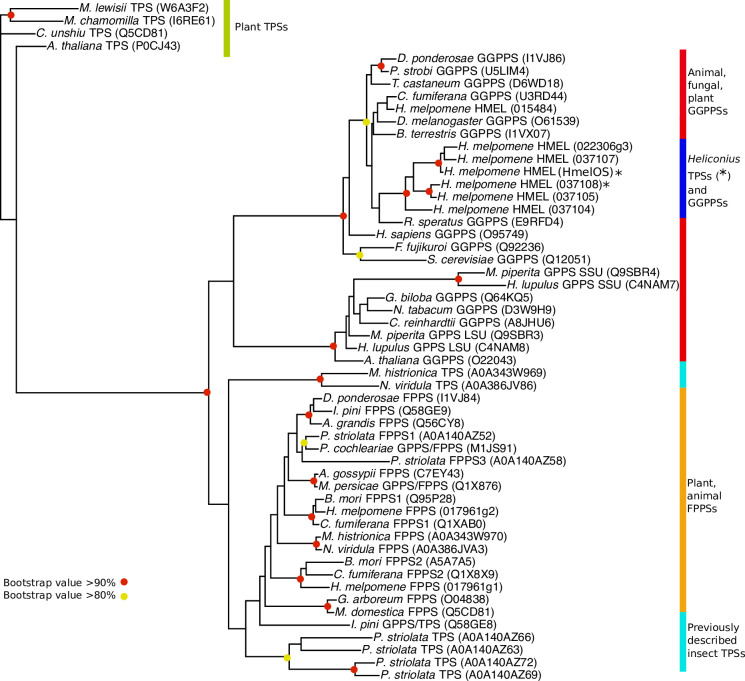
Plants and insects often use the same compounds for chemical communication, but not much is known about the genetics of convergent evolution of chemical signals. The terpene (E)-β-ocimene is a common component of floral scent and is also used by the butterfly Heliconius melpomene as an anti-aphrodisiac pheromone. While the biosynthesis of terpenes has been described in plants and microorganisms, few terpene synthases (TPSs) have been identified in insects. Here, we study the recent divergence of 2 species, H. melpomene and Heliconius cydno, which differ in the presence of (E)-β-ocimene; combining linkage mapping, gene expression, and functional analyses, we identify 2 novel TPSs. Furthermore, we demonstrate that one, HmelOS, is able to synthesise (E)-β-ocimene in vitro. We find no evidence for TPS activity in HcydOS (HmelOS ortholog of H. cydno), suggesting that the loss of (E)-β-ocimene in this species is the result of coding, not regulatory, differences. The TPS enzymes we discovered are unrelated to previously described plant and insect TPSs, demonstrating that chemical convergence has independent evolutionary origins.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
References
-
- Baker TC. Origin of courtship and sex pheromones of the oriental fruit moth and a discussion of the role of phytochemicals in the evolution of lepidopteran male scents. In: Chou CH, Waller GR, editors. Phytochemical Ecology: Allelochemicals, Mycotoxins, and Insect Pheromones and Allomones. Taipei, Taiwan; 1989. pp. 401–418.
-
- Conner WE, Iyengar VK. Male pheromones in moths: Reproductive isolation, sexy sons, and good genes In: Allison JD, Carde RT, editors. Pheromone Communication in Moths: Evolution, Behavior, and Application. Berkeley: University of California Press; 2016. pp. 191–208. 10.1525/9780520964433-013 - DOI
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
