

T-bet-dependent ILC1- and NK cell-derived IFN-γ mediates cDC1-dependent host resistance against Toxoplasma gondii
- PMID: 33465134
- PMCID: PMC7875365
- DOI: 10.1371/journal.ppat.1008299
T-bet-dependent ILC1- and NK cell-derived IFN-γ mediates cDC1-dependent host resistance against Toxoplasma gondii
Abstract
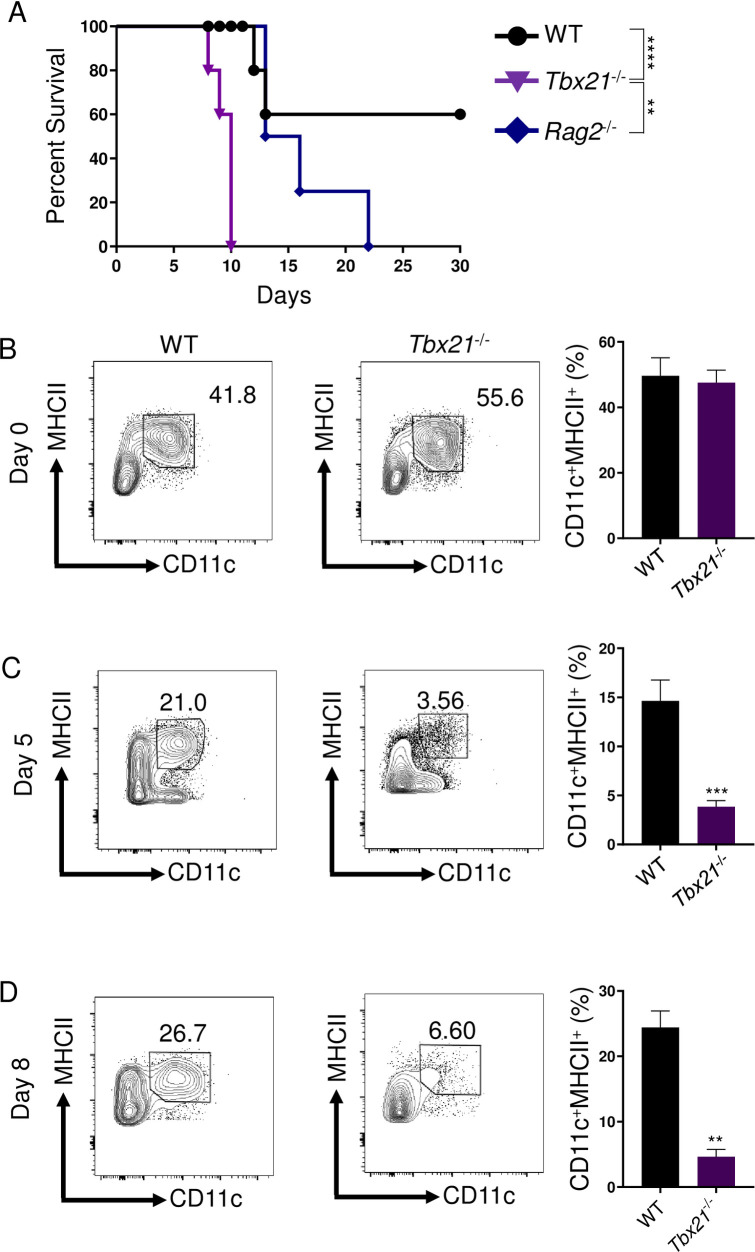
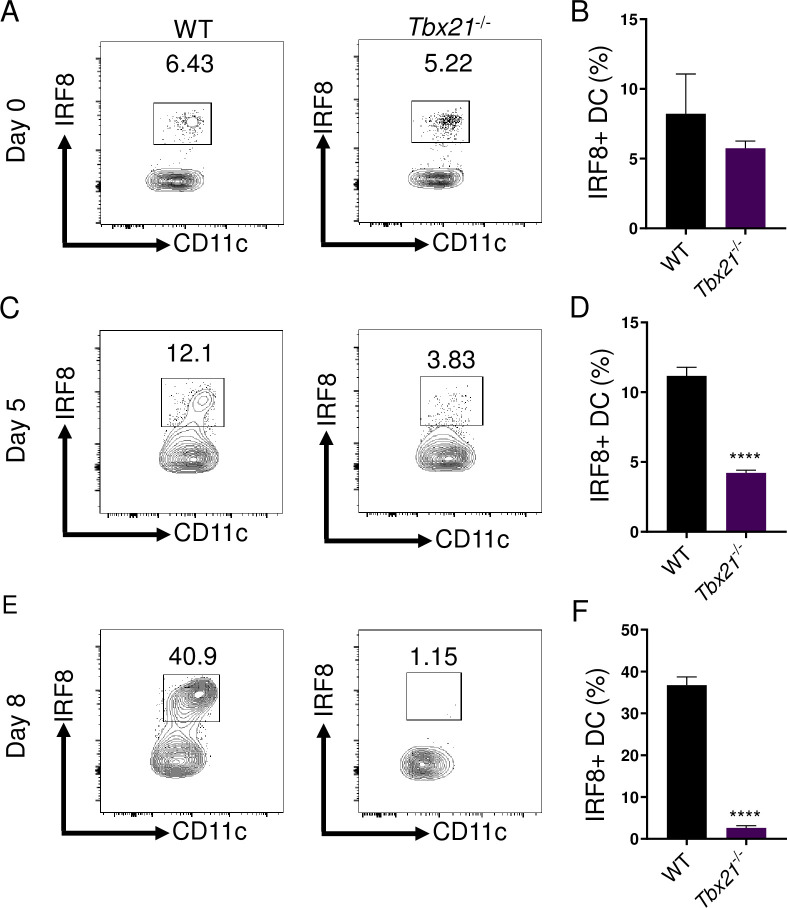
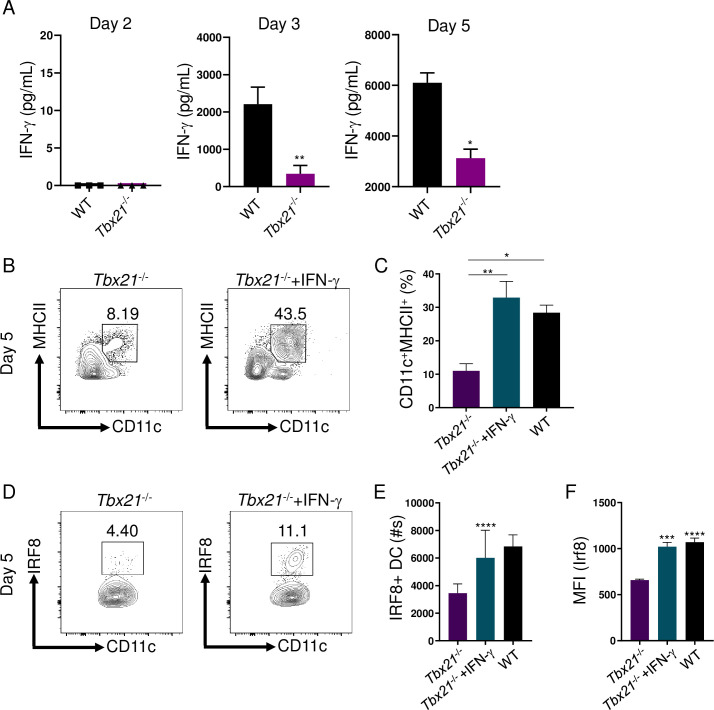
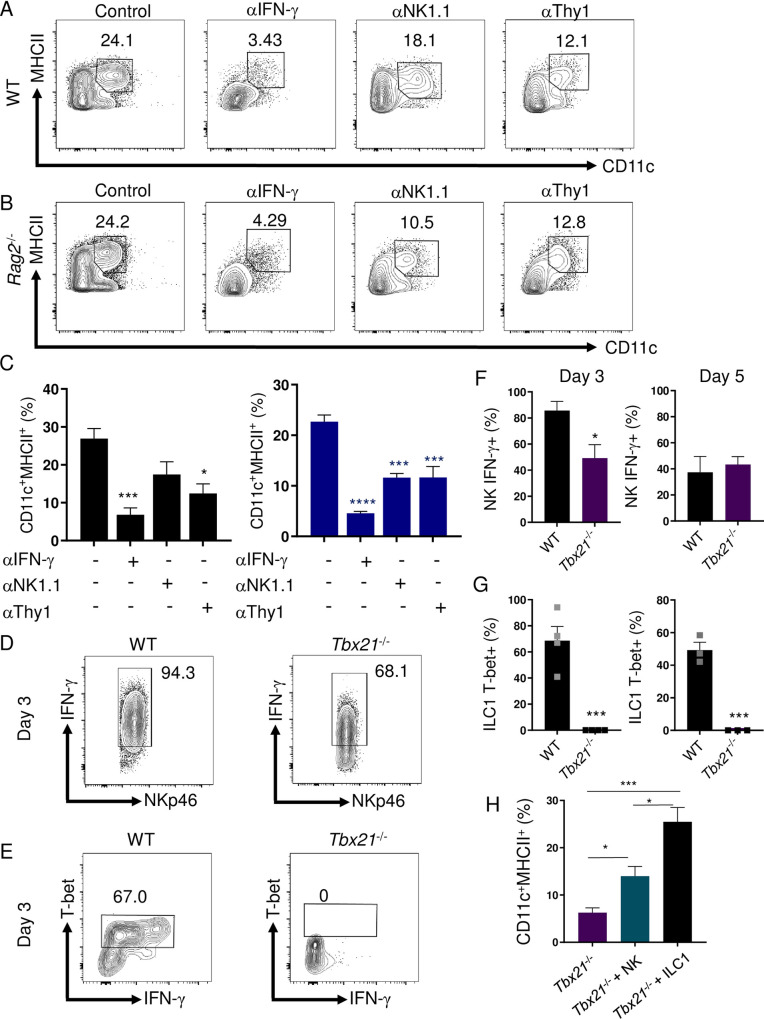
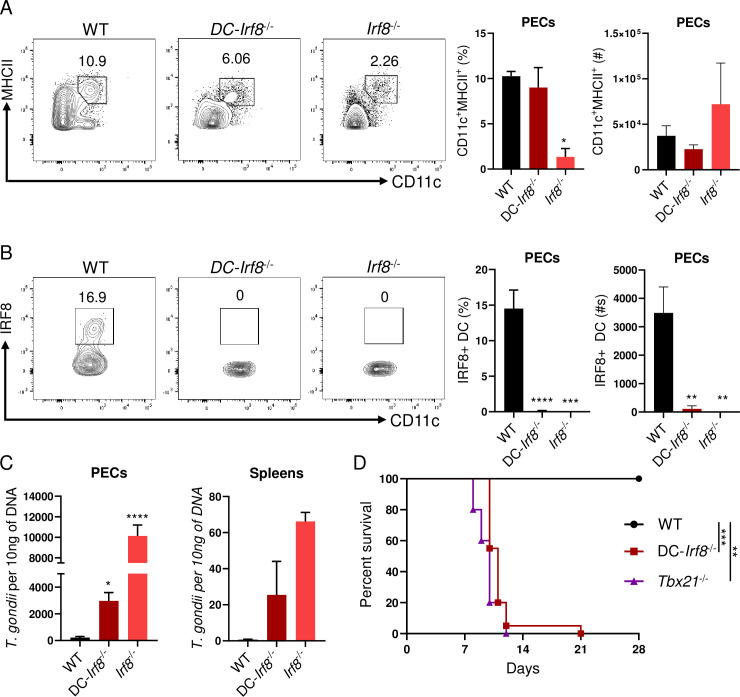
Host resistance against intracellular pathogens requires a rapid IFN-γ mediated immune response. We reveal that T-bet-dependent production of IFN-γ is essential for the maintenance of inflammatory DCs at the site of infection with a common protozoan parasite, Toxoplasma gondii. A detailed analysis of the cellular sources for T-bet-dependent IFN-γ identified that ILC1s and to a lesser degree NK, but not TH1 cells, were involved in the regulation of inflammatory DCs via IFN-γ. Mechanistically, we established that T-bet dependent innate IFN-γ is critical for the induction of IRF8, an essential transcription factor for cDC1s. Failure to upregulate IRF8 in DCs resulted in acute susceptibility to T. gondii infection. Our data identifies that T-bet dependent production of IFN-γ by ILC1 and NK cells is indispensable for host resistance against intracellular infection via maintaining IRF8+ inflammatory DCs at the site of infection.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
Similar articles
-
IL-12 Mediates T-bet-Expressing Myeloid Cell-Dependent Host Resistance against Toxoplasma gondii.Immunohorizons. 2024 Apr 1;8(4):355-362. doi: 10.4049/immunohorizons.2400029. Immunohorizons. 2024. PMID: 38687282 Free PMC article.
-
The cat's out of the bag: Toxoplasma gondii provides further insight into myeloid-mediated host defense.Immunohorizons. 2025 Aug 25;9(9):vlaf037. doi: 10.1093/immhor/vlaf037. Immunohorizons. 2025. PMID: 40854593 Free PMC article. Review.
-
C5aR1 Activation Drives Early IFN-γ Production to Control Experimental Toxoplasma gondii Infection.Front Immunol. 2020 Jul 8;11:1397. doi: 10.3389/fimmu.2020.01397. eCollection 2020. Front Immunol. 2020. PMID: 32733463 Free PMC article.
-
IL-33 promotes innate lymphoid cell-dependent IFN-γ production required for innate immunity to Toxoplasma gondii.Elife. 2021 Apr 30;10:e65614. doi: 10.7554/eLife.65614. Elife. 2021. PMID: 33929319 Free PMC article.
-
Induction and regulation of IL-12-dependent host resistance to Toxoplasma gondii.Immunol Res. 2003;27(2-3):521-8. doi: 10.1385/IR:27:2-3:521. Immunol Res. 2003. PMID: 12857995 Review.
Cited by
-
The role of IFN-γ-mediated host immune responses in monitoring and the elimination of Toxoplasma gondii infection.Int Immunol. 2024 Apr 3;36(5):199-210. doi: 10.1093/intimm/dxae001. Int Immunol. 2024. PMID: 38175650 Free PMC article. Review.
-
Impact of MyD88, Microbiota, and Location on Type 1 and Type 3 Innate Lymphoid Cells during Toxoplasma gondii Infection.Immunohorizons. 2022 Sep 12;6(9):660-670. doi: 10.4049/immunohorizons.2200070. Immunohorizons. 2022. PMID: 36096673 Free PMC article.
-
ILC1: Development, maturation, and transcriptional regulation.Eur J Immunol. 2023 Feb;53(2):e2149435. doi: 10.1002/eji.202149435. Epub 2022 Nov 27. Eur J Immunol. 2023. PMID: 36408791 Free PMC article. Review.
-
Innate Lymphoid Cells in Response to Intracellular Pathogens: Protection Versus Immunopathology.Front Cell Infect Microbiol. 2021 Dec 6;11:775554. doi: 10.3389/fcimb.2021.775554. eCollection 2021. Front Cell Infect Microbiol. 2021. PMID: 34938670 Free PMC article. Review.
-
Immune modulation of Th1/Th2/Treg/Th17/Th9/Th21 cells in rabbits infected with Eimeria stiedai.Front Cell Infect Microbiol. 2023 Aug 1;13:1230689. doi: 10.3389/fcimb.2023.1230689. eCollection 2023. Front Cell Infect Microbiol. 2023. PMID: 37593762 Free PMC article.
References
-
- Sullivan BM, Jobe O, Lazarevic V, Vasquez K, Bronson R, Glimcher LH, et al. Increased susceptibility of mice lacking T-bet to infection with Mycobacterium tuberculosis correlates with increased IL-10 and decreased IFN-gamma production. Journal of immunology. 2005;175(7):4593–602. Epub 2005/09/24. 10.4049/jimmunol.175.7.4593 . - DOI - PubMed
-
- López-Yglesias AH, Burger E, Araujo A, Martin AT, Yarovinsky F. T-bet-independent Th1 response induces intestinal immunopathology during Toxoplasma gondii infection. Mucosal immunology. 2018. 10.1038/mi.2017.102 https://www.nature.com/articles/mi2017102#supplementary-information. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
