

Immunological design of commensal communities to treat intestinal infection and inflammation
- PMID: 33465156
- PMCID: PMC7846104
- DOI: 10.1371/journal.ppat.1009191
Immunological design of commensal communities to treat intestinal infection and inflammation
Abstract
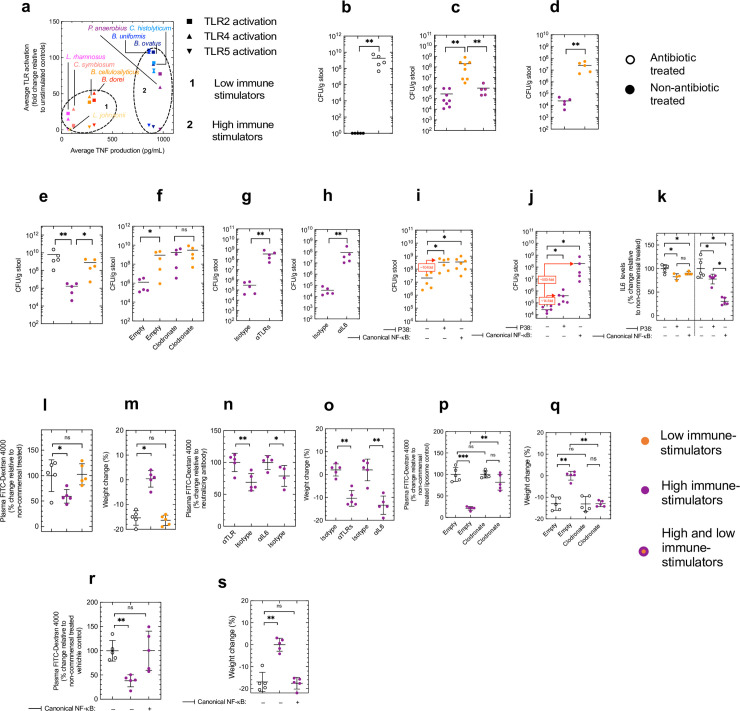
The immunological impact of individual commensal species within the microbiota is poorly understood limiting the use of commensals to treat disease. Here, we systematically profile the immunological fingerprint of commensals from the major phyla in the human intestine (Actinobacteria, Bacteroidetes, Firmicutes and Proteobacteria) to reveal taxonomic patterns in immune activation and use this information to rationally design commensal communities to enhance antibacterial defenses and combat intestinal inflammation. We reveal that Bacteroidetes and Firmicutes have distinct effects on intestinal immunity by differentially inducing primary and secondary response genes. Within these phyla, the immunostimulatory capacity of commensals from the Bacteroidia class (Bacteroidetes phyla) reflects their robustness of TLR4 activation and Bacteroidia communities rely solely on this receptor for their effects on intestinal immunity. By contrast, within the Clostridia class (Firmicutes phyla) it reflects the degree of TLR2 and TLR4 activation, and communities of Clostridia signal via both of these receptors to exert their effects on intestinal immunity. By analyzing the receptors, intracellular signaling components and transcription factors that are engaged by different commensal species, we identify canonical NF-κB signaling as a critical rheostat which grades the degree of immune stimulation commensals elicit. Guided by this immunological analysis, we constructed a cross-phylum consortium of commensals (Bacteroides uniformis, Bacteroides ovatus, Peptostreptococcus anaerobius and Clostridium histolyticum) which enhances innate TLR, IL6 and macrophages-dependent defenses against intestinal colonization by vancomycin resistant Enterococci, and fortifies mucosal barrier function during pathological intestinal inflammation through the same pathway. Critically, the setpoint of intestinal immunity established by this consortium is calibrated by canonical NF-κB signaling. Thus, by profiling the immunological impact of major human commensal species our work paves the way for rational microbiota reengineering to protect against antibiotic resistant infections and to treat intestinal inflammation.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures



Similar articles
-
The Roles of Inflammation, Nutrient Availability and the Commensal Microbiota in Enteric Pathogen Infection.Microbiol Spectr. 2015 Jun;3(3). doi: 10.1128/microbiolspec.MBP-0008-2014. Microbiol Spectr. 2015. PMID: 26185088
-
Gnotobiotic IL-10-/-;NF-kappa B(EGFP) mice reveal the critical role of TLR/NF-kappa B signaling in commensal bacteria-induced colitis.J Immunol. 2007 May 15;178(10):6522-32. doi: 10.4049/jimmunol.178.10.6522. J Immunol. 2007. PMID: 17475882
-
Innate and adaptive immunity cooperate flexibly to maintain host-microbiota mutualism.Science. 2009 Jul 31;325(5940):617-20. doi: 10.1126/science.1172747. Science. 2009. PMID: 19644121 Free PMC article.
-
Bacterial colonization and the development of intestinal defences.Can J Gastroenterol. 2004 Aug;18(8):493-500. doi: 10.1155/2004/690421. Can J Gastroenterol. 2004. PMID: 15372112 Review.
-
Interaction Between Commensal Bacteria, Immune Response and the Intestinal Barrier in Inflammatory Bowel Disease.Front Immunol. 2021 Nov 11;12:761981. doi: 10.3389/fimmu.2021.761981. eCollection 2021. Front Immunol. 2021. PMID: 34858414 Free PMC article. Review.
Cited by
-
Analysis of Cow-Calf Microbiome Transfer Routes and Microbiome Diversity in the Newborn Holstein Dairy Calf Hindgut.Front Nutr. 2021 Oct 25;8:736270. doi: 10.3389/fnut.2021.736270. eCollection 2021. Front Nutr. 2021. PMID: 34760909 Free PMC article.
-
Peptostreptococcus Anaerobius enhances dextran sulfate sodium-induced colitis by promoting nf-κB-NLRP3-Dependent macrophage pyroptosis.Virulence. 2024 Dec;15(1):2435391. doi: 10.1080/21505594.2024.2435391. Epub 2024 Nov 29. Virulence. 2024. PMID: 39611567 Free PMC article.
-
The Potential of Gut Microbiota in Prediction of Stroke-Associated Pneumonia.Brain Sci. 2023 Aug 17;13(8):1217. doi: 10.3390/brainsci13081217. Brain Sci. 2023. PMID: 37626573 Free PMC article.
-
Maternal diet and gut microbiome composition modulate early-life immune development.EMBO Mol Med. 2023 Aug 7;15(8):e17241. doi: 10.15252/emmm.202217241. Epub 2023 Jun 6. EMBO Mol Med. 2023. PMID: 37278126 Free PMC article.
-
The gut microbial metabolic capacity of microbiome-humanized vs. wild type rodents reveals a likely dual role of intestinal bacteria in hepato-intestinal schistosomiasis.PLoS Negl Trop Dis. 2022 Oct 24;16(10):e0010878. doi: 10.1371/journal.pntd.0010878. eCollection 2022 Oct. PLoS Negl Trop Dis. 2022. PMID: 36279280 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
