

Activated intestinal muscle cells promote preadipocyte migration: a novel mechanism for creeping fat formation in Crohn's disease
- PMID: 33468536
- PMCID: PMC8286985
- DOI: 10.1136/gutjnl-2020-323719
Activated intestinal muscle cells promote preadipocyte migration: a novel mechanism for creeping fat formation in Crohn's disease
Abstract
Objective: Creeping fat, the wrapping of mesenteric fat around the bowel wall, is a typical feature of Crohn's disease, and is associated with stricture formation and bowel obstruction. How creeping fat forms is unknown, and we interrogated potential mechanisms using novel intestinal tissue and cell interaction systems.
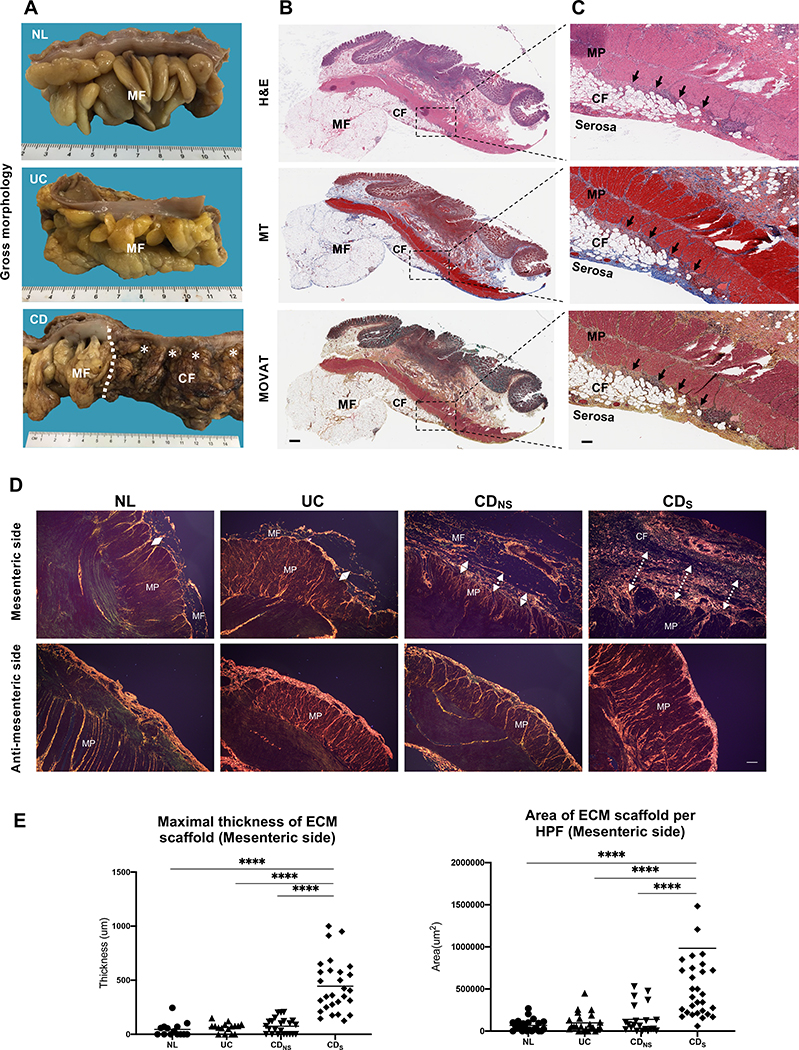
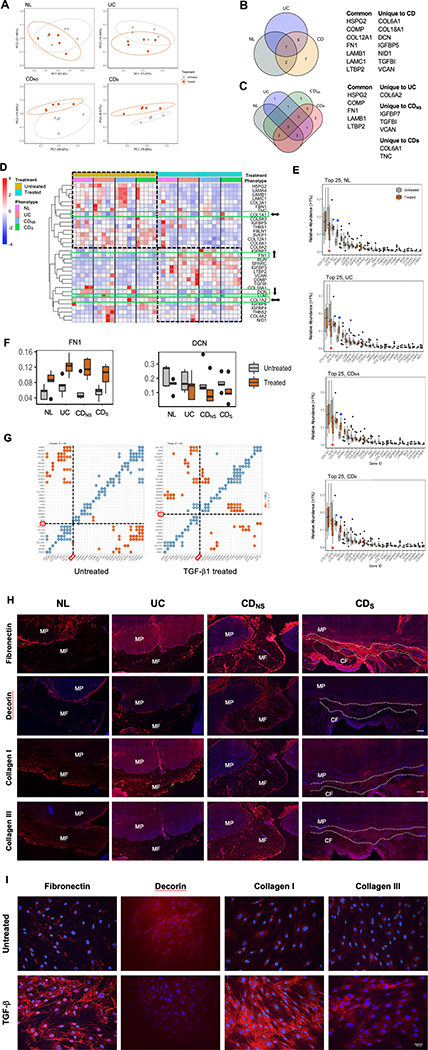
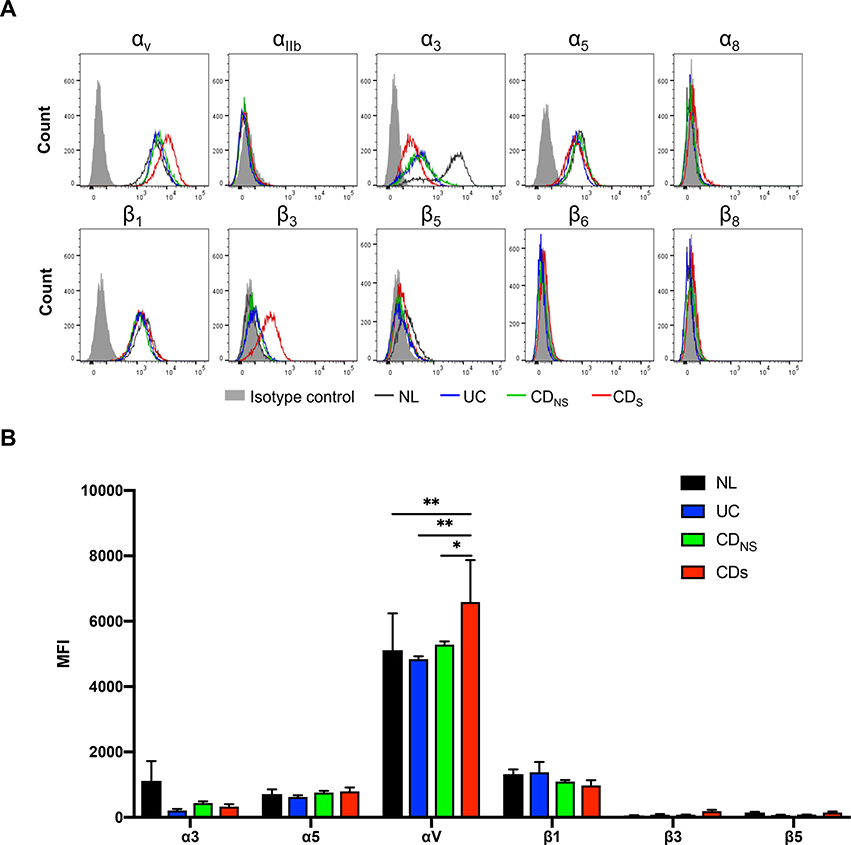
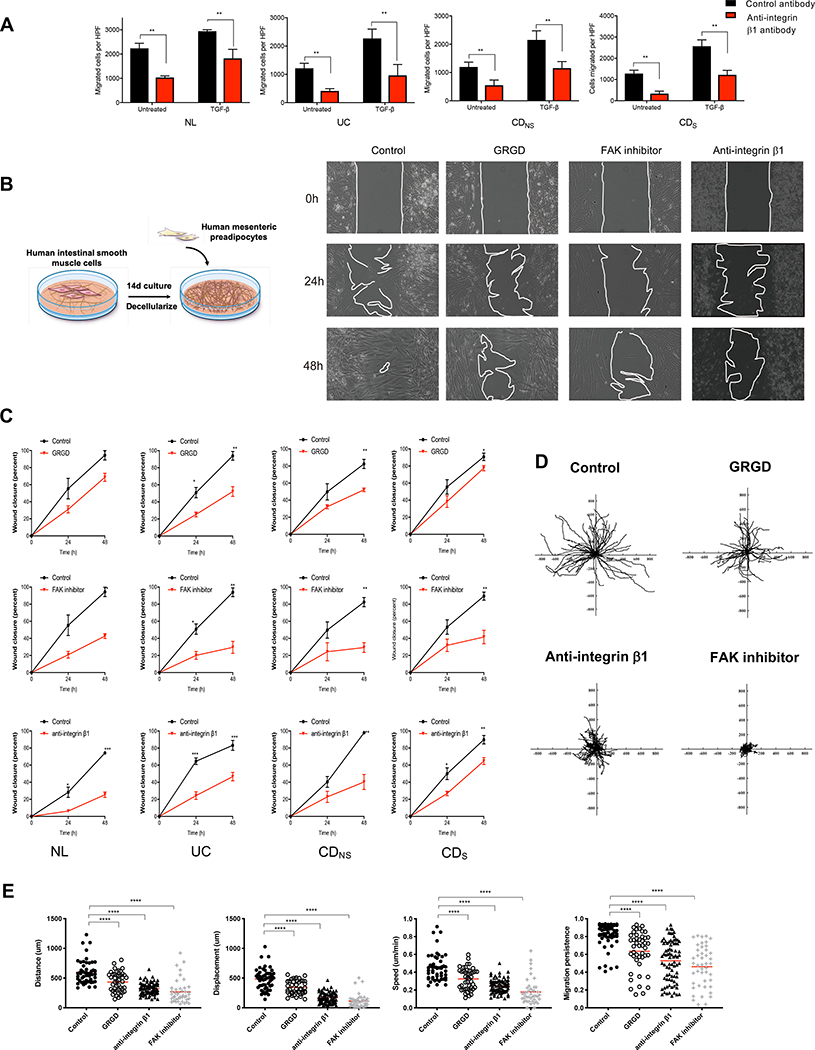
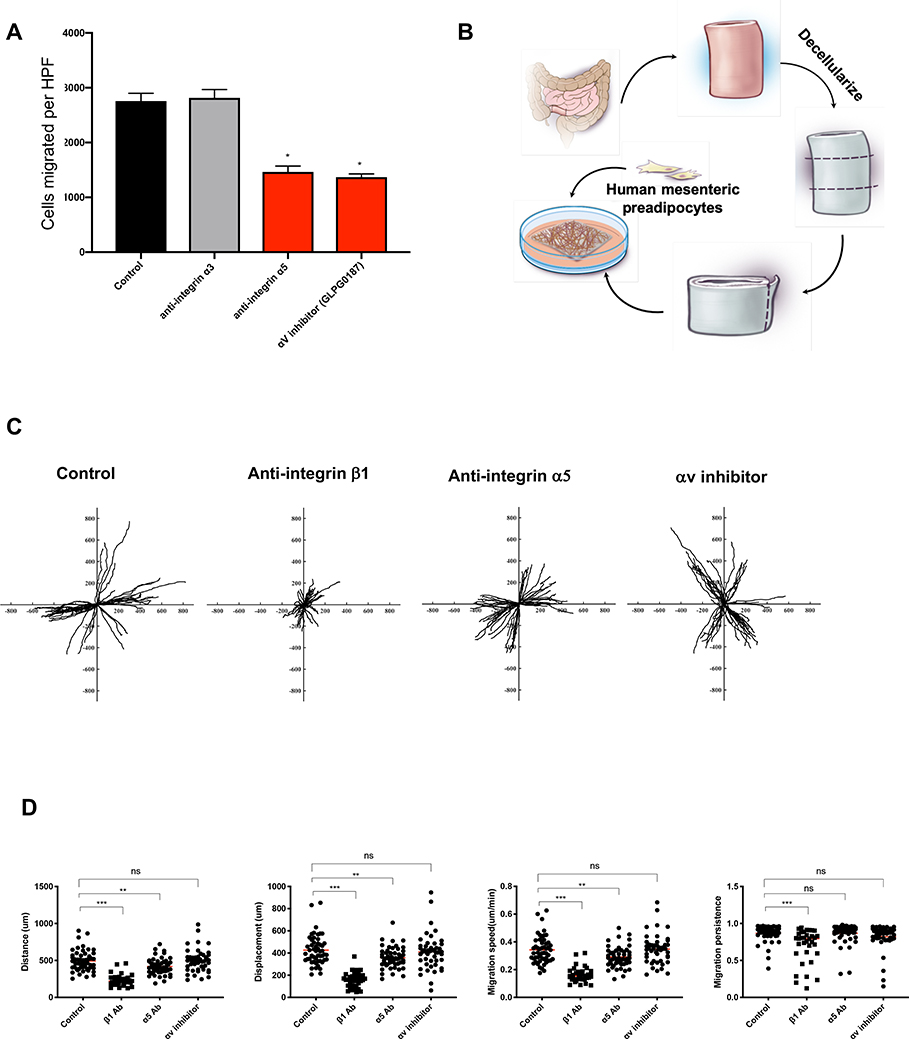
Design: Tissues from normal, UC, non-strictured and strictured Crohn's disease intestinal specimens were obtained. The muscularis propria matrisome was determined via proteomics. Mesenteric fat explants, primary human preadipocytes and adipocytes were used in multiple ex vivo and in vitro cell migration systems on muscularis propria muscle cell derived or native extracellular matrix. Functional experiments included integrin characterisation via flow cytometry and their inhibition with specific blocking antibodies and chemicals.
Results: Crohn's disease muscularis propria cells produced an extracellular matrix scaffold which is in direct spatial and functional contact with the immediately overlaid creeping fat. The scaffold contained multiple proteins, but only fibronectin production was singularly upregulated by transforming growth factor-β1. The muscle cell-derived matrix triggered migration of preadipocytes out of mesenteric fat, fibronectin being the dominant factor responsible for their migration. Blockade of α5β1 on the preadipocyte surface inhibited their migration out of mesenteric fat and on 3D decellularised intestinal tissue extracellular matrix.
Conclusion: Crohn's disease creeping fat appears to result from the migration of preadipocytes out of mesenteric fat and differentiation into adipocytes in response to an increased production of fibronectin by activated muscularis propria cells. These new mechanistic insights may lead to novel approaches for prevention of creeping fat-associated stricture formation.
Keywords: Crohn's disease; extracellular matrix; fibrosis.
© Author(s) (or their employer(s)) 2022. No commercial re-use. See rights and permissions. Published by BMJ.
Conflict of interest statement
Competing interests: FR is consultant to Agomab, Allergan, AbbVie, Boehringer-Ingelheim, Celgene, Cowen, Genentech, Gilead, Gossamer, Guidepoint, Helmsley, Index Pharma, Jannsen, Koutif, Metacrine, Morphic, Pfizer, Pliant, Prometheus Biosciences, Receptos, RedX, Roche, Samsung, Takeda, Techlab, Thetis, UCB and receives funding from the Crohn’s and Colitis Foundation of America, the Helmsley Charitable Trust, Kenneth Rainin Foundation and the National Institute of Health. CF received speaker fees from UCB, Genentech, Sandoz, Janssen and he is consultant for Athos Therapeutics.
Figures


References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical