

The Phosphatidyl- myo-Inositol Dimannoside Acyltransferase PatA Is Essential for Mycobacterium tuberculosis Growth In Vitro and In Vivo
- PMID: 33468587
- PMCID: PMC8088522
- DOI: 10.1128/JB.00439-20
The Phosphatidyl- myo-Inositol Dimannoside Acyltransferase PatA Is Essential for Mycobacterium tuberculosis Growth In Vitro and In Vivo
Abstract
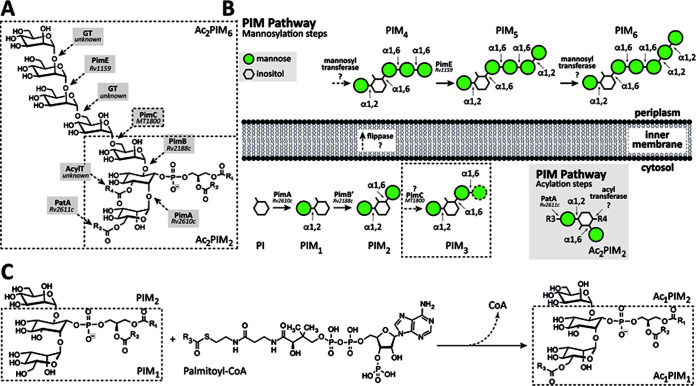
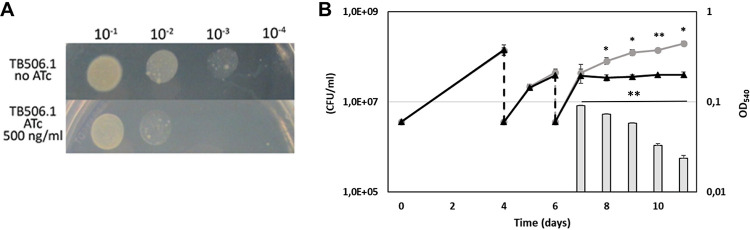
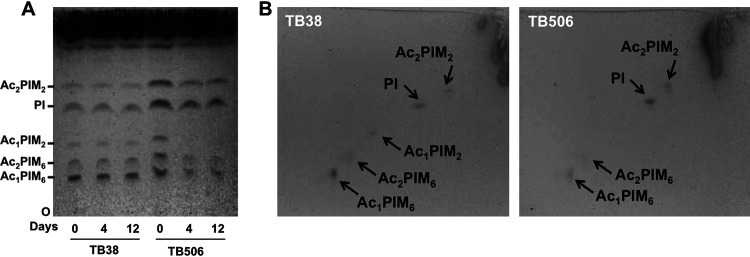
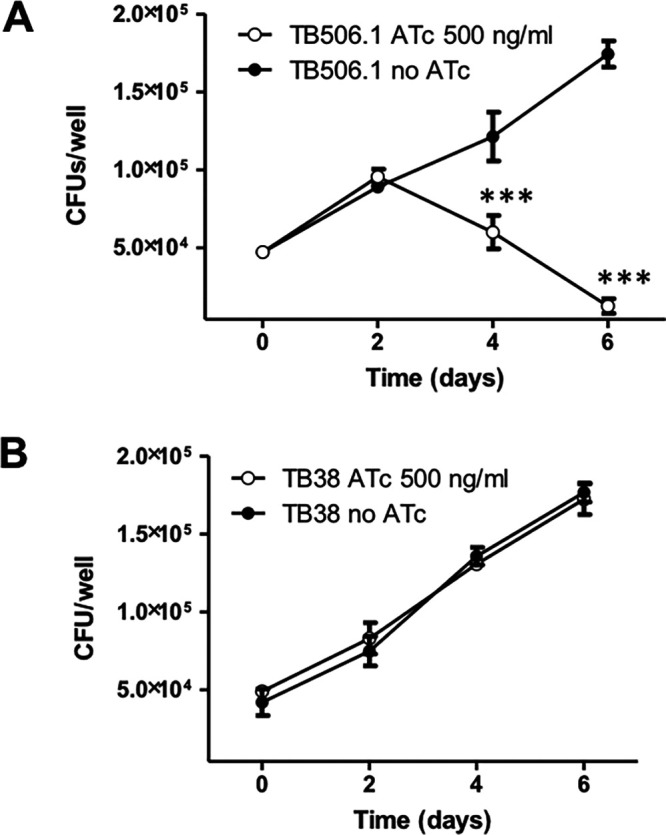
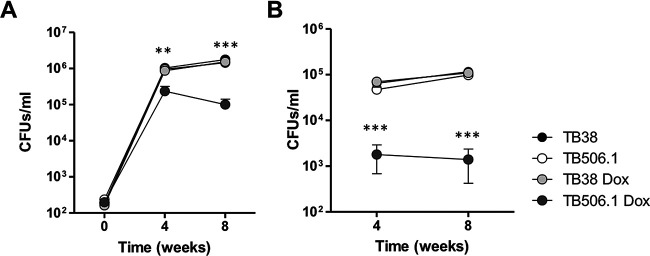
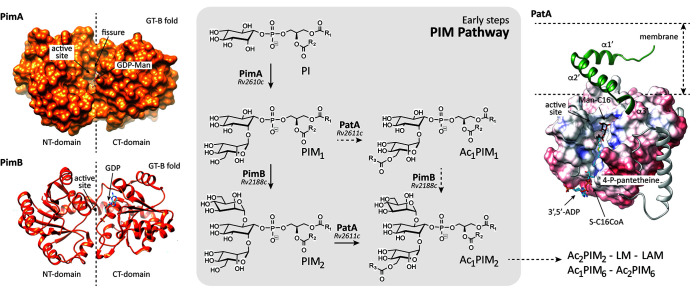
Mycobacterium tuberculosis comprises an unusual cell envelope dominated by unique lipids and glycans that provides a permeability barrier against hydrophilic drugs and is central for its survival and virulence. Phosphatidyl-myo-inositol mannosides (PIMs) are glycolipids considered to be not only key structural components of the cell envelope but also the precursors of lipomannan (LM) and lipoarabinomannan (LAM), important lipoglycans implicated in host-pathogen interactions. Here, we focus on PatA, a membrane-associated acyltransferase that transfers a palmitoyl moiety from palmitoyl coenzyme A (palmitoyl-CoA) to the 6-position of the mannose ring linked to the 2-position of inositol in PIM1/PIM2 We validate that the function of PatA is vital for M. tuberculosisin vitro and in vivo We constructed a patA conditional mutant and showed that silencing patA is bactericidal in batch cultures. This phenotype was associated with significantly reduced levels of Ac1PIM2, an important structural component of the mycobacterial inner membrane. The requirement of PatA for viability was also demonstrated during macrophage infection and in a mouse model of infection, where a dramatic decrease in viable counts was observed upon silencing of the patA gene. This is reminiscent of the behavior of PimA, the mannosyltransferase that initiates the PIM pathway, also found to be essential for M. tuberculosis growth in vitro and in vivo Altogether, the experimental data highlight the significance of the early steps of the PIM biosynthetic pathway for M. tuberculosis physiology and reveal that PatA is a novel target for drug discovery programs against this major human pathogen.IMPORTANCE Tuberculosis (TB) is the leading cause of death from a single infectious agent. The emergence of drug resistance in strains of M. tuberculosis, the etiologic agent of TB, emphasizes the need to identify new targets and antimicrobial agents. The mycobacterial cell envelope is a major factor in this intrinsic drug resistance. Here, we have focused on the biosynthesis of PIMs, key virulence factors and important components of the cell envelope. Specifically, we have determined that PatA, the acyltransferase responsible for the first acylation step of the PIM synthesis pathway, is essential in M. tuberculosis These results highlight the importance of early steps of the PIM biosynthetic pathway for mycobacterial physiology and the suitability of PatA as a potential new drug target.
Keywords: Mycobacterium tuberculosis; acyltransferase; conditional mutant; glycolipid; mycobacterium; tuberculosis.
Copyright © 2021 American Society for Microbiology.
Figures
References
-
- World Health Organization. 2019. Global tuberculosis report 2019. World Health Organization, Geneva, Switzerland.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
