

De Novo Cobalamin Biosynthesis, Transport, and Assimilation and Cobalamin-Mediated Regulation of Methionine Biosynthesis in Mycobacterium smegmatis
- PMID: 33468593
- PMCID: PMC8088520
- DOI: 10.1128/JB.00620-20
De Novo Cobalamin Biosynthesis, Transport, and Assimilation and Cobalamin-Mediated Regulation of Methionine Biosynthesis in Mycobacterium smegmatis
Erratum in
-
Correction for Kipkorir et al., "De Novo Cobalamin Biosynthesis, Transport, and Assimilation and Cobalamin-Mediated Regulation of Methionine Biosynthesis in Mycobacterium smegmatis".J Bacteriol. 2022 Mar 15;204(3):e0000422. doi: 10.1128/jb.00004-22. Epub 2022 Mar 15. J Bacteriol. 2022. PMID: 35289639 Free PMC article. No abstract available.
Abstract
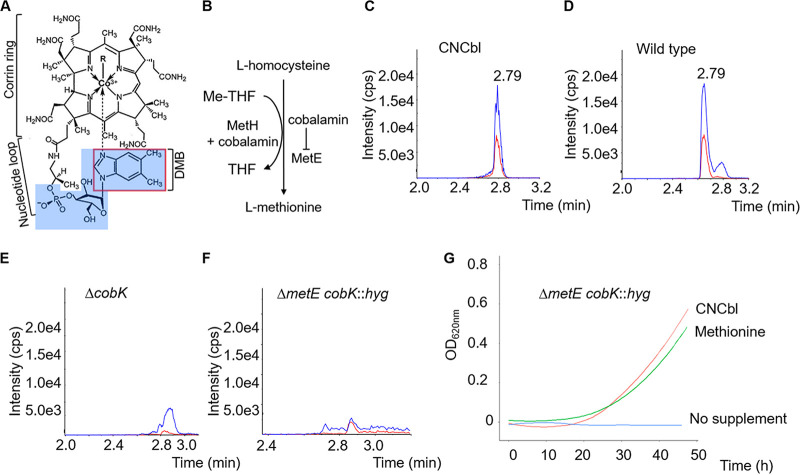
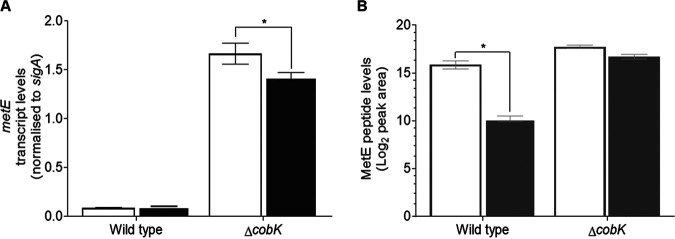
Cobalamin is an essential cofactor in all domains of life, yet its biosynthesis is restricted to some bacteria and archaea. Mycobacterium smegmatis, an environmental saprophyte frequently used as surrogate for the obligate human pathogen M. tuberculosis, carries approximately 30 genes predicted to be involved in de novo cobalamin biosynthesis. M. smegmatis also encodes multiple cobalamin-dependent enzymes, including MetH, a methionine synthase that catalyzes the final reaction in methionine biosynthesis. In addition to metH, M. smegmatis possesses a cobalamin-independent methionine synthase, metE, suggesting that enzyme use-MetH versus MetE-is regulated by cobalamin availability. Consistent with this notion, we previously described a cobalamin-sensing riboswitch controlling metE expression in M. tuberculosis Here, we apply a targeted mass spectrometry-based approach to confirm de novo cobalamin biosynthesis in M. smegmatis during aerobic growth in vitro We also demonstrate that M. smegmatis can transport and assimilate exogenous cyanocobalamin (CNCbl; also known as vitamin B12) and its precursor, dicyanocobinamide ([CN]2Cbi). However, the uptake of CNCbl and (CN)2Cbi in this organism is restricted and seems dependent on the conditional essentiality of the cobalamin-dependent methionine synthase. Using gene and protein expression analyses combined with single-cell growth kinetics and live-cell time-lapse microscopy, we show that transcription and translation of metE are strongly attenuated by endogenous cobalamin. These results support the inference that metH essentiality in M. smegmatis results from riboswitch-mediated repression of MetE expression. Moreover, differences observed in cobalamin-dependent metabolism between M. smegmatis and M. tuberculosis provide some insight into the selective pressures which might have shaped mycobacterial metabolism for pathogenicity.IMPORTANCE Alterations in cobalamin-dependent metabolism have marked the evolution of Mycobacterium tuberculosis into a human pathogen. However, the role(s) of cobalamin in mycobacterial physiology remains poorly understood. Using the nonpathogenic saprophyte M. smegmatis, we investigated the production of cobalamin, transport and assimilation of cobalamin precursors, and the role of cobalamin in regulating methionine biosynthesis. We confirm constitutive de novo cobalamin biosynthesis in M. smegmatis, in contrast with M. tuberculosis, which appears to lack de novo cobalamin biosynthetic capacity. We also show that uptake of cyanocobalamin (vitamin B12) and its precursors is restricted in M. smegmatis, apparently depending on the cofactor requirements of the cobalamin-dependent methionine synthase. These observations establish M. smegmatis as an informative foil to elucidate key metabolic adaptations enabling mycobacterial pathogenicity.
Keywords: Mycobacterium tuberculosis; cobK; mycobacterial metabolism; riboswitch; tuberculosis; vitamin B12.
Copyright © 2021 Kipkorir et al.
Figures




References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
