

Fungal foraging behaviour and hyphal space exploration in micro-structured Soil Chips
- PMID: 33469165
- PMCID: PMC8163874
- DOI: 10.1038/s41396-020-00886-7
Fungal foraging behaviour and hyphal space exploration in micro-structured Soil Chips
Abstract
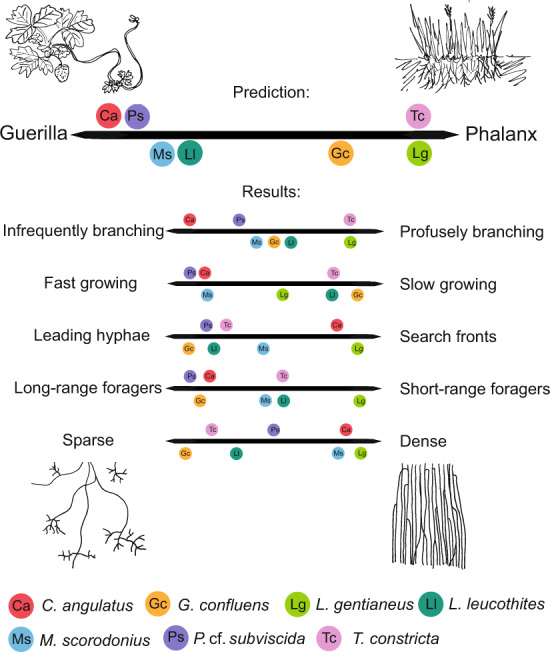
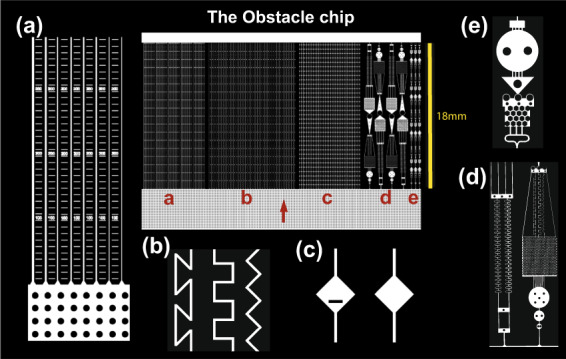
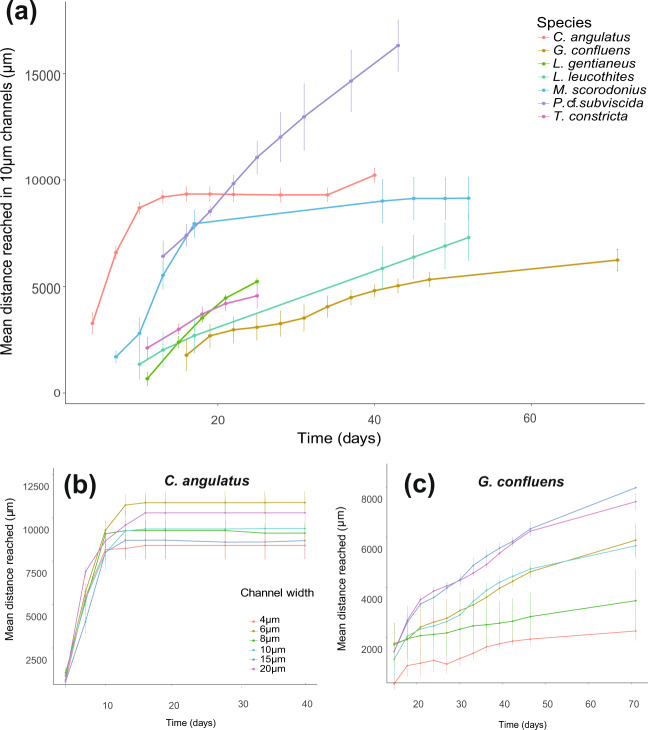
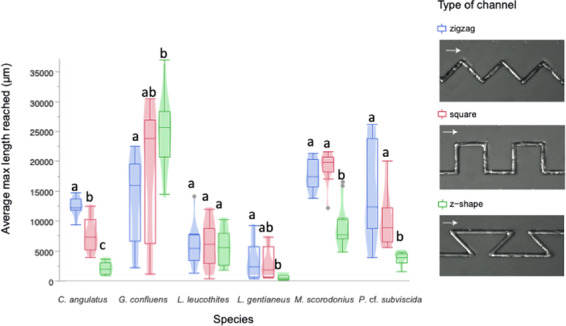
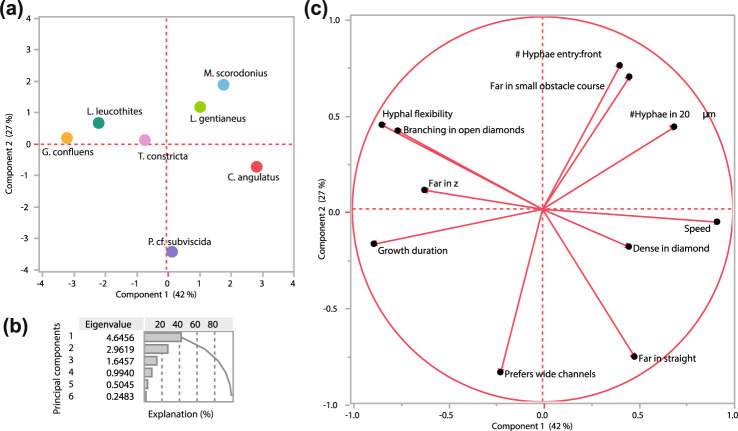
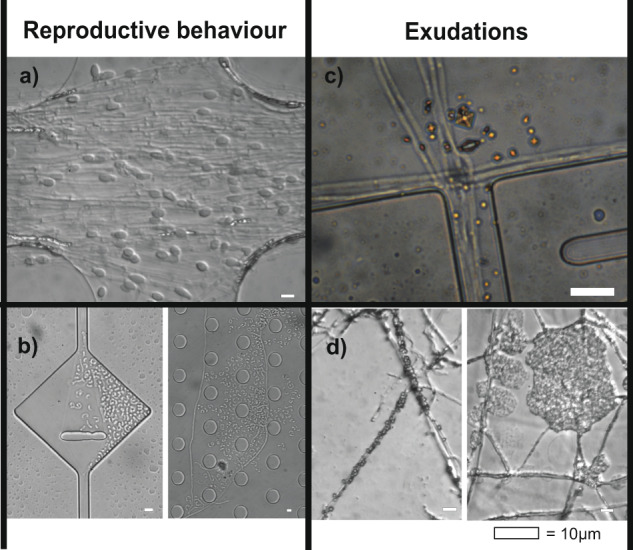
How do fungi navigate through the complex microscopic maze-like structures found in the soil? Fungal behaviour, especially at the hyphal scale, is largely unknown and challenging to study in natural habitats such as the opaque soil matrix. We monitored hyphal growth behaviour and strategies of seven Basidiomycete litter decomposing species in a micro-fabricated "Soil Chip" system that simulates principal aspects of the soil pore space and its micro-spatial heterogeneity. The hyphae were faced with micrometre constrictions, sharp turns and protruding obstacles, and the species examined were found to have profoundly different responses in terms of foraging range and persistence, spatial exploration and ability to pass obstacles. Hyphal behaviour was not predictable solely based on ecological assumptions, and our results obtained a level of trait information at the hyphal scale that cannot be fully explained using classical concepts of space exploration and exploitation such as the phalanx/guerrilla strategies. Instead, we propose a multivariate trait analysis, acknowledging the complex trade-offs and microscale strategies that fungal mycelia exhibit. Our results provide novel insights about hyphal behaviour, as well as an additional understanding of fungal habitat colonisation, their foraging strategies and niche partitioning in the soil environment.
Conflict of interest statement
The authors declare that they have no conflict of interest.
Figures


References
-
- Ferguson BA, Dreisbach TA, Parks CG, Filip GM, Schmitt CL. Coarse-scale population structure of pathogenic Armillaria species in a mixed-conifer forest in the Blue Mountains of northeast Oregon. Can J Res. 2003;33:612–23.
-
- Smith SE, Read DJ. Mycorrhizal symbiosis. 3rd ed. London, UK: Academic Press; 2008.
-
- Maron JL, Marler M, Klironomos JN, Cleveland CC. Soil fungal pathogens and the relationship between plant diversity and productivity. Ecol Lett. 2011;14:36–41. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
