

Long-Term Exercise Reduces Formation of Tubular Aggregates and Promotes Maintenance of Ca2+ Entry Units in Aged Muscle
- PMID: 33469430
- PMCID: PMC7813885
- DOI: 10.3389/fphys.2020.601057
Long-Term Exercise Reduces Formation of Tubular Aggregates and Promotes Maintenance of Ca2+ Entry Units in Aged Muscle
Abstract
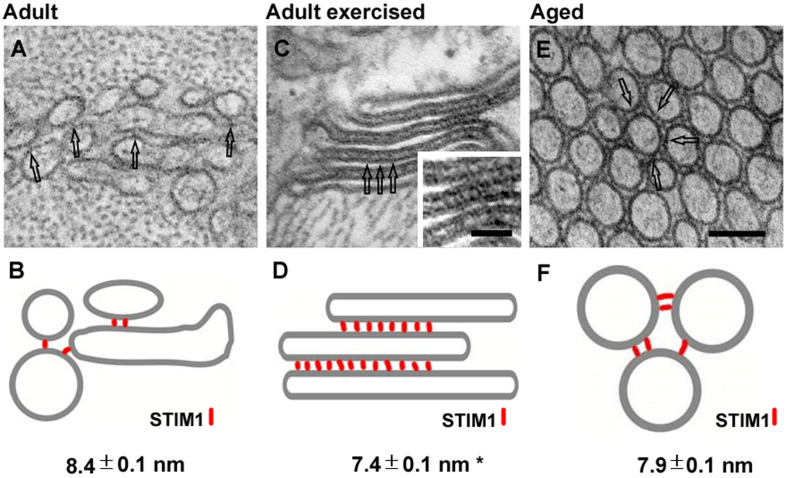
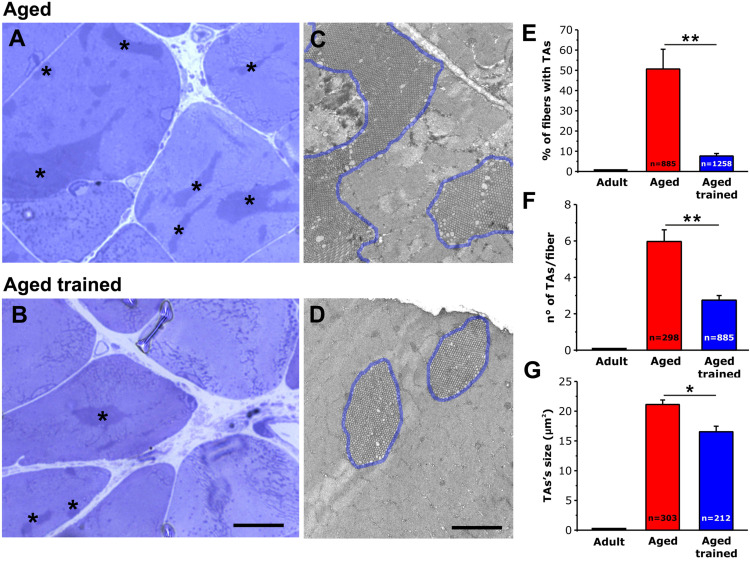
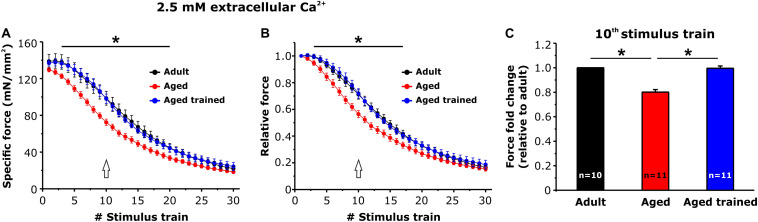
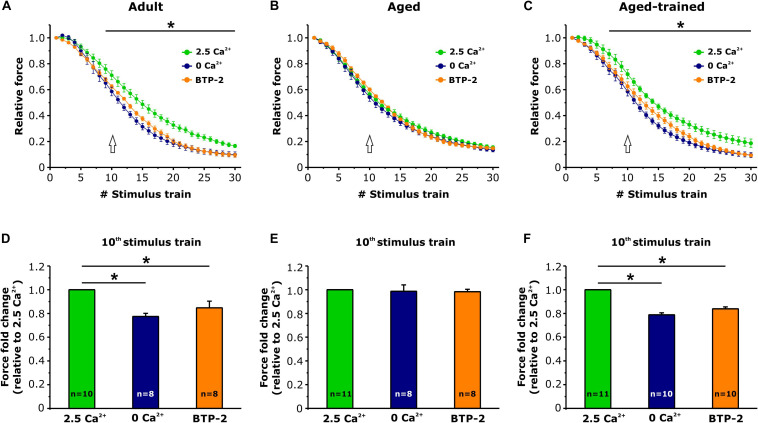
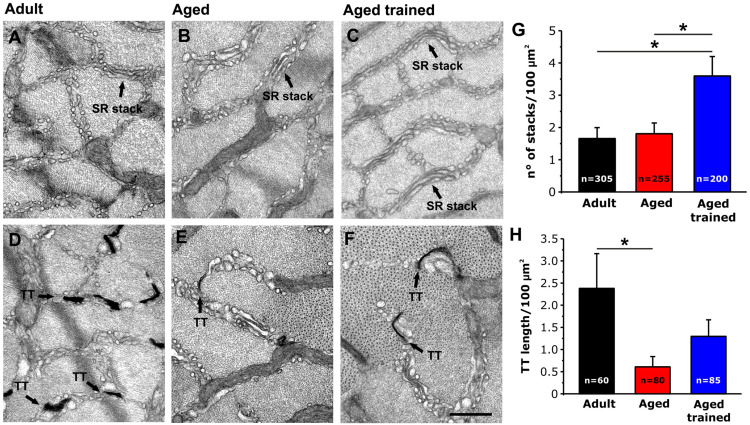
Tubular aggregates (TAs) in skeletal muscle fibers are unusual accumulation of sarcoplasmic reticulum (SR) tubes that are found in different disorders including TA myopathy (TAM). TAM is a muscular disease characterized by muscle pain, cramping, and weakness that has been recently linked to mutations in STIM1 and ORAI1. STIM1 and ORAI1 are the two main proteins mediating store-operated Ca2+ entry (SOCE), a mechanism activated by depletion of intracellular Ca2+ stores (e.g., SR) that allows recovery of Ca2+ from the extracellular space during repetitive muscle activity. We have recently shown that exercise triggers the formation of unique intracellular junctions between SR and transverse tubules named Ca 2+ entry units (CEUs). CEUs promote colocalization of STIM1 with ORAI1 and improve muscle function in presence of external Ca2+. TAs virtually identical to those of TAM patients are also found in fast-twitch fibers of aging male mice. Here, we used a combination of electron and confocal microscopy, Western blotting, and ex vivo stimulation protocols (in presence or absence of external Ca2+) to evaluate the presence of TAs, STIM1-ORAI1 localization and expression and fatigue resistance of intact extensor digitorum longus (EDL) muscles in wild-type male adult (4-month-old) and aged (24-month-old) mice and in mice trained in wheel cages for 15 months (from 9 to 24 months of age). The results collected indicate that (i) aging causes STIM1 and ORAI1 to accumulate in TAs and (ii) long-term exercise significantly reduced formation of TAs. In addition, (iii) EDL muscles from aged mice exhibited a faster decay of contractile force than adult muscles, likely caused by their inability to refill intracellular Ca2+ stores, and (iv) exercise in wheel cages restored the capability of aged EDL muscles to use external Ca2+ by promoting maintenance of CEUs. In conclusion, exercise prevented improper accumulation of STIM1 and ORAI1 in TAs during aging, maintaining the capability of aged muscle to refill intracellular Ca2+ stores via SOCE.
Keywords: electron microscopy; excitation-contraction coupling; sarcoplasmic reticulum; store opereted calcium entry; transverse tubule.
Copyright © 2021 Boncompagni, Pecorai, Michelucci, Pietrangelo and Protasi.
Conflict of interest statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Figures

Comment in
-
Commentary: Long-Term Exercise Reduces Formation of Tubular Aggregates and Promotes Maintenance of Ca2+ Entry Units in Aged Muscle.Front Physiol. 2021 Apr 1;12:663677. doi: 10.3389/fphys.2021.663677. eCollection 2021. Front Physiol. 2021. PMID: 33868028 Free PMC article. No abstract available.
Similar articles
-
Constitutive, Muscle-Specific Orai1 Knockout Results in the Incomplete Assembly of Ca2+ Entry Units and a Reduction in the Age-Dependent Formation of Tubular Aggregates.Biomedicines. 2024 Jul 24;12(8):1651. doi: 10.3390/biomedicines12081651. Biomedicines. 2024. PMID: 39200116 Free PMC article.
-
Calcium entry units (CEUs): perspectives in skeletal muscle function and disease.J Muscle Res Cell Motil. 2021 Jun;42(2):233-249. doi: 10.1007/s10974-020-09586-3. Epub 2020 Aug 18. J Muscle Res Cell Motil. 2021. PMID: 32812118 Free PMC article.
-
Searching for Mechanisms Underlying the Assembly of Calcium Entry Units: The Role of Temperature and pH.Int J Mol Sci. 2023 Mar 10;24(6):5328. doi: 10.3390/ijms24065328. Int J Mol Sci. 2023. PMID: 36982401 Free PMC article.
-
Role of STIM1/ORAI1-mediated store-operated Ca2+ entry in skeletal muscle physiology and disease.Cell Calcium. 2018 Dec;76:101-115. doi: 10.1016/j.ceca.2018.10.004. Epub 2018 Oct 30. Cell Calcium. 2018. PMID: 30414508 Free PMC article. Review.
-
Store-operated calcium entry: From physiology to tubular aggregate myopathy.Curr Opin Pharmacol. 2023 Feb;68:102347. doi: 10.1016/j.coph.2022.102347. Epub 2023 Jan 4. Curr Opin Pharmacol. 2023. PMID: 36608411 Review.
Cited by
-
Commentary: Long-Term Exercise Reduces Formation of Tubular Aggregates and Promotes Maintenance of Ca2+ Entry Units in Aged Muscle.Front Physiol. 2021 Apr 1;12:663677. doi: 10.3389/fphys.2021.663677. eCollection 2021. Front Physiol. 2021. PMID: 33868028 Free PMC article. No abstract available.
-
The Effects of Aging on Sarcoplasmic Reticulum-Related Factors in the Skeletal Muscle of Mice.Int J Mol Sci. 2024 Feb 10;25(4):2148. doi: 10.3390/ijms25042148. Int J Mol Sci. 2024. PMID: 38396828 Free PMC article.
-
Role of store-operated Ca2+ entry in cardiovascular disease.Cell Commun Signal. 2022 Mar 18;20(1):33. doi: 10.1186/s12964-022-00829-z. Cell Commun Signal. 2022. PMID: 35303866 Free PMC article. Review.
-
Assessing the Potential of Nutraceuticals as Geroprotectors on Muscle Performance and Cognition in Aging Mice.Antioxidants (Basel). 2021 Sep 4;10(9):1415. doi: 10.3390/antiox10091415. Antioxidants (Basel). 2021. PMID: 34573047 Free PMC article.
-
Constitutive, Muscle-Specific Orai1 Knockout Results in the Incomplete Assembly of Ca2+ Entry Units and a Reduction in the Age-Dependent Formation of Tubular Aggregates.Biomedicines. 2024 Jul 24;12(8):1651. doi: 10.3390/biomedicines12081651. Biomedicines. 2024. PMID: 39200116 Free PMC article.
References
-
- Boncompagni S., d’Amelio L., Fulle S., Fanò G., Protasi F. (2006). Progressive disorganization of the excitation-contraction coupling apparatus in aging human skeletal muscle as revealed by electron microscopy: a possible role in the decline of muscle performance. J. Gerontol. A Biol. Sci. Med. Sci. 61 995–1008. 10.1093/gerona/61.10.995 - DOI - PubMed
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous