

Designed PKC-targeting bryostatin analogs modulate innate immunity and neuroinflammation
- PMID: 33472023
- PMCID: PMC8052272
- DOI: 10.1016/j.chembiol.2020.12.015
Designed PKC-targeting bryostatin analogs modulate innate immunity and neuroinflammation
Abstract
Neuroinflammation characterizes multiple neurologic diseases, including primary inflammatory conditions such as multiple sclerosis and classical neurodegenerative diseases. Aberrant activation of the innate immune system contributes to disease progression, but drugs modulating innate immunity, particularly within the central nervous system (CNS), are lacking. The CNS-penetrant natural product bryostatin-1 attenuates neuroinflammation by targeting innate myeloid cells. Supplies of natural bryostatin-1 are limited, but a recent scalable good manufacturing practice (GMP) synthesis has enabled access to it and its analogs (bryologs), the latter providing a path to more efficacious, better tolerated, and more accessible agents. Here, we show that multiple synthetically accessible bryologs replicate the anti-inflammatory effects of bryostatin-1 on innate immune cells in vitro, and a lead bryolog attenuates neuroinflammation in vivo, actions mechanistically dependent on protein kinase C (PKC) binding. Our findings identify bryologs as promising drug candidates for targeting innate immunity in neuroinflammation and create a platform for evaluation of synthetic PKC modulators in neuroinflammatory diseases.
Keywords: EAE; PKC; bryolog; bryostatin; innate immunity; microglia; multiple sclerosis; neuroinflammation; prostratin.
Copyright © 2020 Elsevier Ltd. All rights reserved.
Conflict of interest statement
Declaration of interests Johns Hopkins University and Stanford University have filed patent applications on this and related technology. Stanford University patent applications have been licensed by Neurotrope BioScience for the treatment of neurological disorders and by Bryologyx Inc. for use in HIV/AIDS eradication and cancer immunotherapy. P.A.W. is an advisor to both companies and a cofounder of the latter. M.D.K. has received consulting fees from OptumRx and Biogen Idec. The remaining authors declare no competing interests.
Figures
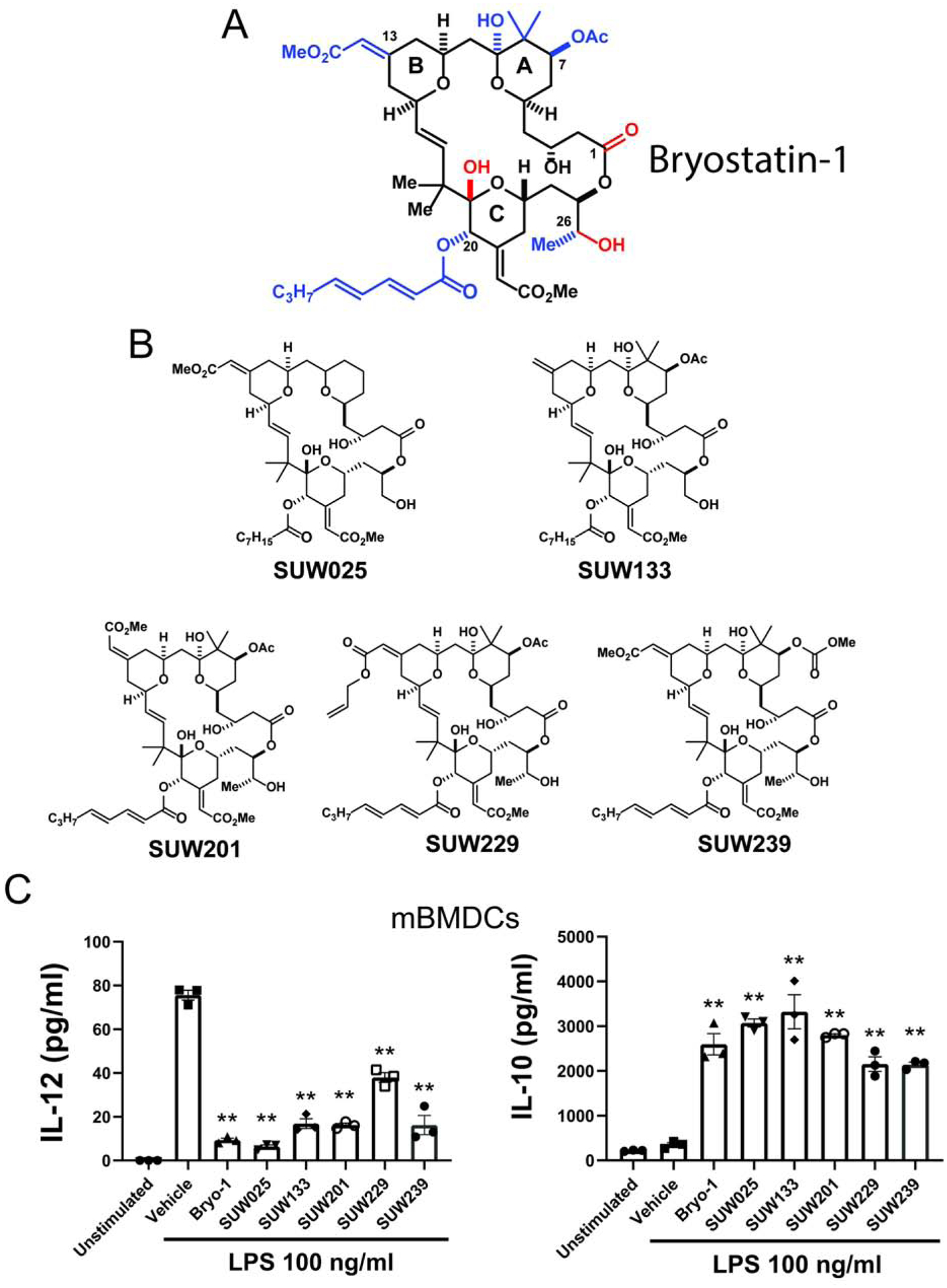


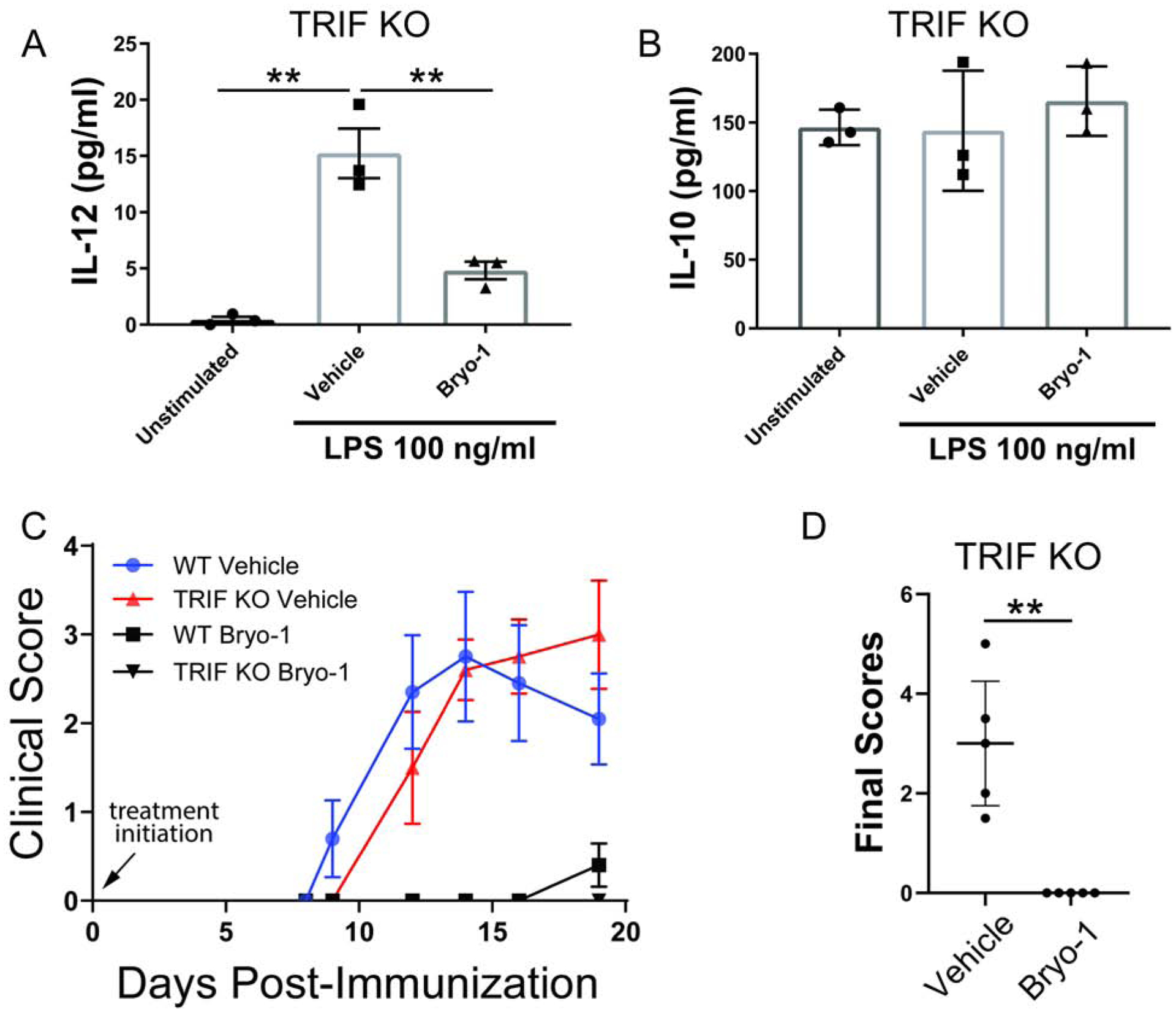

References
-
- Boonstra A, Rajsbaum R, Holman M, Marques R, Asselin-Paturel C, Pereira JP, Bates EEM, Akira S, Vieira P, Liu Y-J, et al. (2006). Macrophages and Myeloid Dendritic Cells, but Not Plasmacytoid Dendritic Cells, Produce IL-10 in Response to MyD88- and TRIF-Dependent TLR Signals, and TLR-Independent Signals. J. Immunol 177, 7551–7558. - PubMed
-
- Clamp A and Jayson GC (2002). The clinical development of the bryostatins. Anticancer Drugs 13, 673–683. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
