

Acute stress drives global repression through two independent RNA polymerase II stalling events in Saccharomyces
- PMID: 33472084
- PMCID: PMC7879390
- DOI: 10.1016/j.celrep.2020.108640
Acute stress drives global repression through two independent RNA polymerase II stalling events in Saccharomyces
Abstract
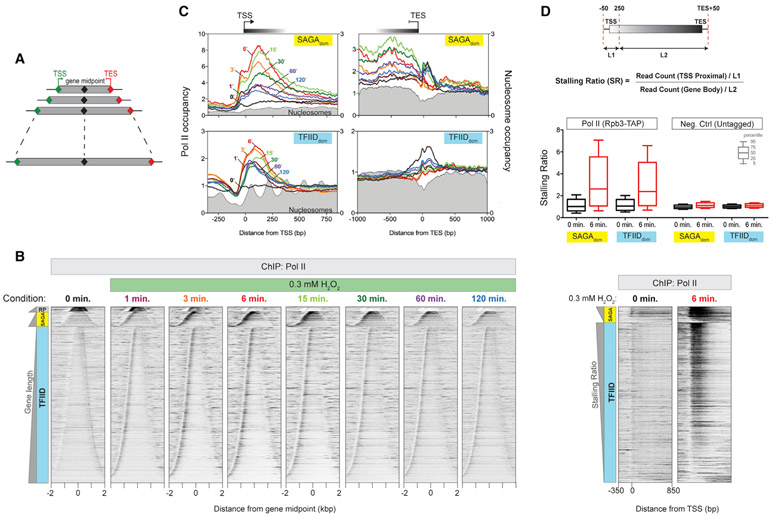
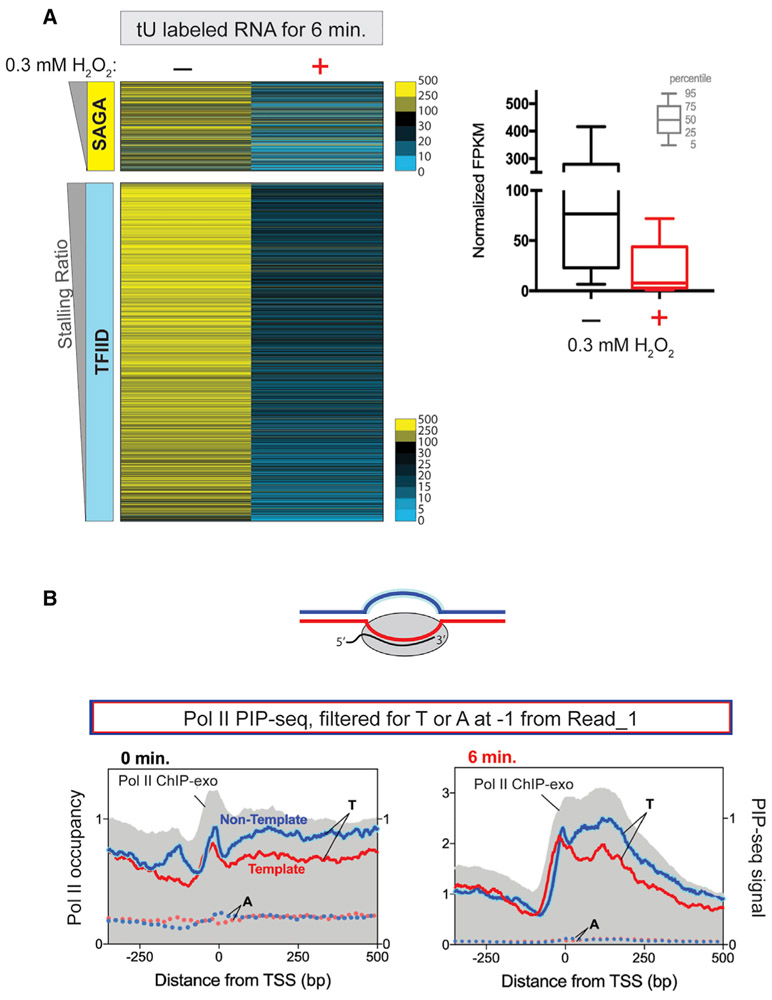
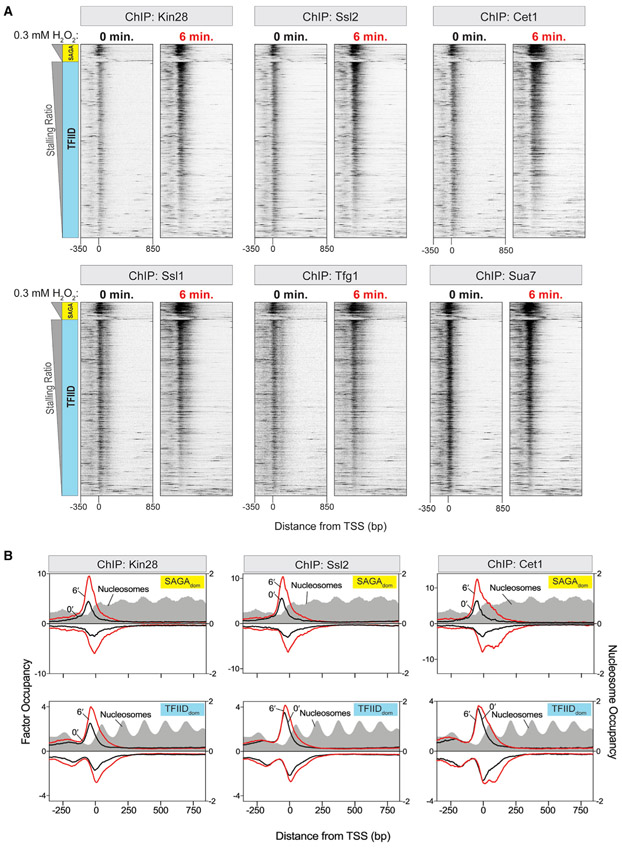
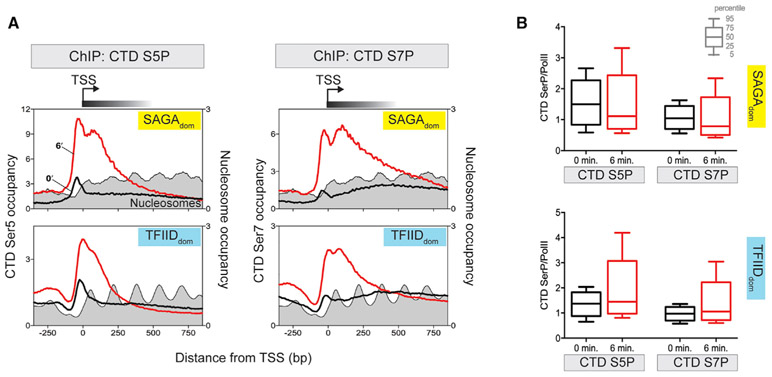
In multicellular eukaryotes, RNA polymerase (Pol) II pauses transcription ~30-50 bp after initiation. While the budding yeast Saccharomyces has its transcription mechanisms mostly conserved with other eukaryotes, it appears to lack this fundamental promoter-proximal pausing. However, we now report that nearly all yeast genes, including constitutive and inducible genes, manifest two distinct transcriptional stall sites that are brought on by acute environmental signaling (e.g., peroxide stress). Pol II first stalls at the pre-initiation stage before promoter clearance, but after DNA melting and factor acquisition, and may involve inhibited dephosphorylation. The second stall occurs at the +2 nucleosome. It acquires most, but not all, elongation factor interactions. Its regulation may include Bur1/Spt4/5. Our results suggest that a double Pol II stall is a mechanism to downregulate essentially all genes in concert.
Keywords: DSIF; Pol II pausing; peroxide stress; promoter-proximal pausing; transcription elongation.
Copyright © 2020 The Author(s). Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests B.F.P. is an owner of and has a financial interest in Peconic, which uses the ChIP-exo technology (U.S. Patent 20100323361A1) implemented in this study and could potentially benefit from the outcomes of this research. All other authors declare no competing interests.
Figures
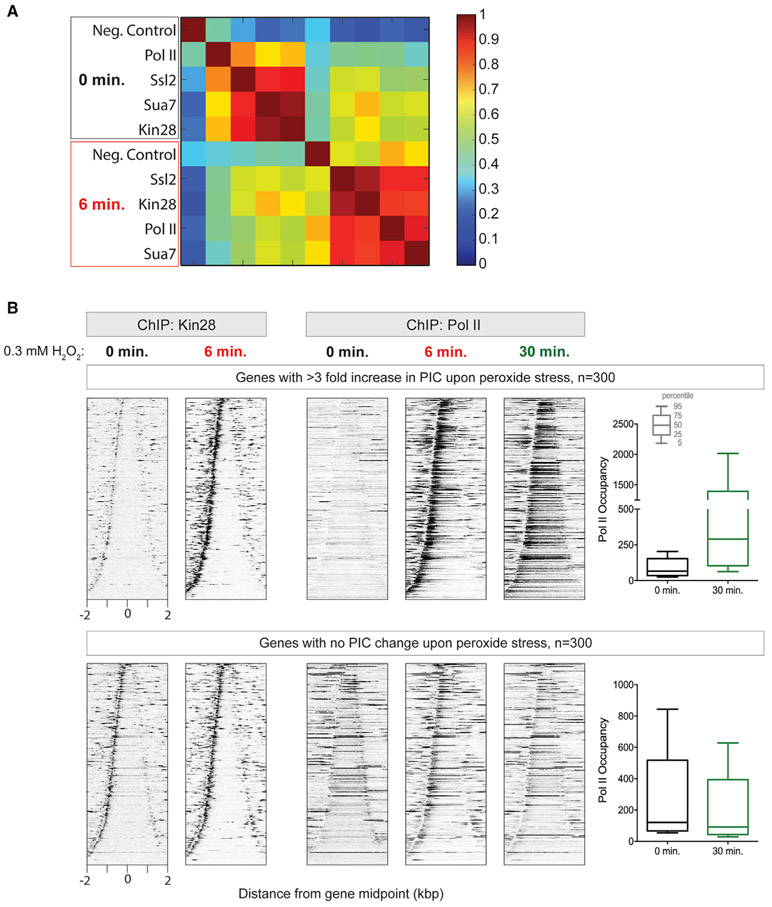
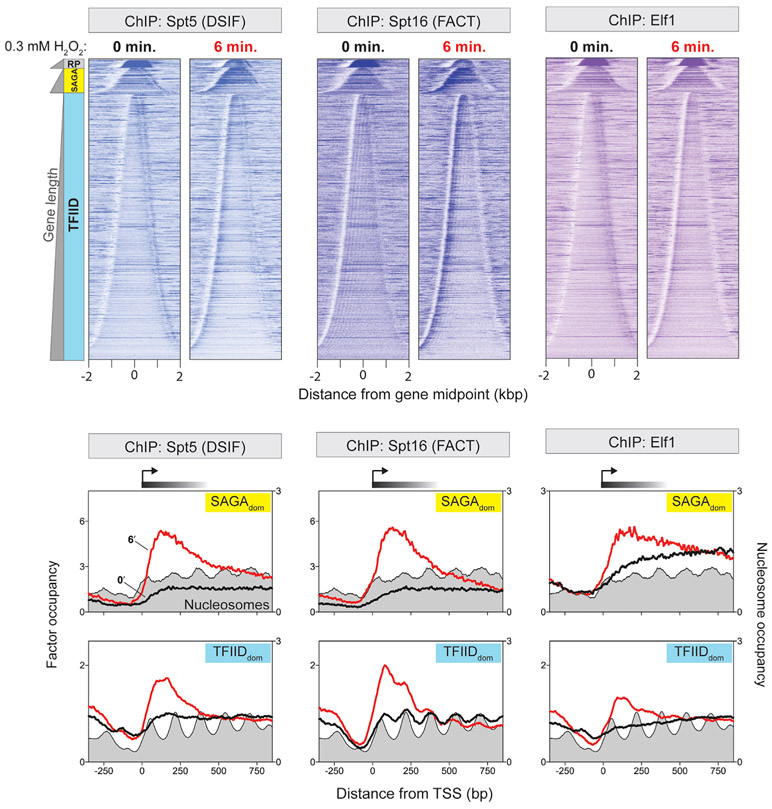
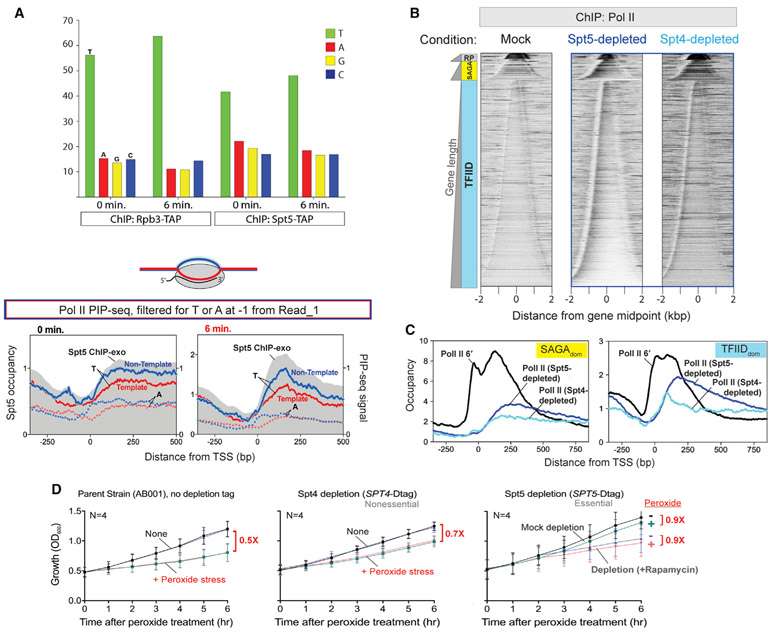
References
-
- Akoulitchev S, Mäkelä TP, Weinberg RA, and Reinberg D (1995). Requirement for TFIIH kinase activity in transcription by RNA polymerase II. Nature 377, 557–560. - PubMed
-
- Allen TA, Von Kaenel S, Goodrich JA, and Kugel JF (2004). The SINE-encoded mouse B2 RNA represses mRNA transcription in response to heat shock. Nat. Struct. Mol. Biol 11, 816–821. - PubMed
-
- Bataille AR, Jeronimo C, Jacques PE, Laramée L, Fortin ME, Forest A, Bergeron M, Hanes SD, and Robert F (2012).A universal RNA polymerase II CTD cycle is orchestrated by complex interplays between kinase, phosphatase, and isomerase enzymes along genes. Mol. Cell 45, 158–170. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
