

Translational landscape and protein biogenesis demands of the early secretory pathway in Komagataella phaffii
- PMID: 33472617
- PMCID: PMC7816318
- DOI: 10.1186/s12934-020-01489-9
Translational landscape and protein biogenesis demands of the early secretory pathway in Komagataella phaffii
Abstract
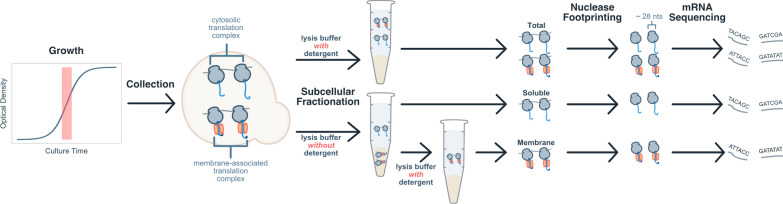
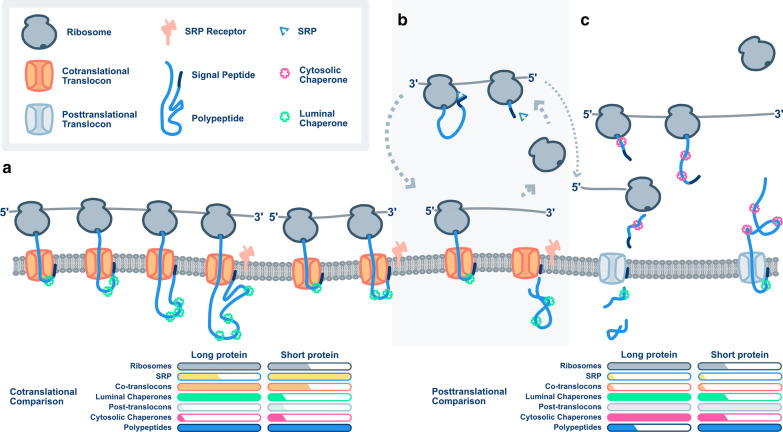
Background: Eukaryotes use distinct networks of biogenesis factors to synthesize, fold, monitor, traffic, and secrete proteins. During heterologous expression, saturation of any of these networks may bottleneck titer and yield. To understand the flux through various routes into the early secretory pathway, we quantified the global and membrane-associated translatomes of Komagataella phaffii.
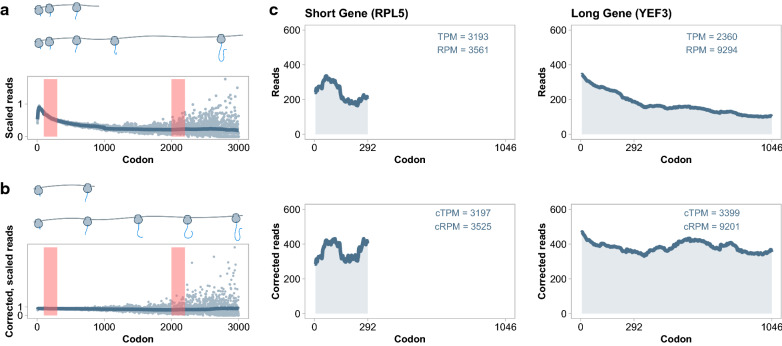
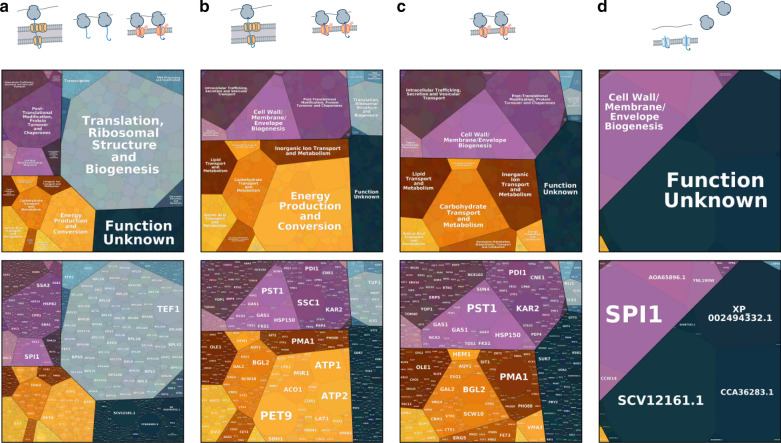
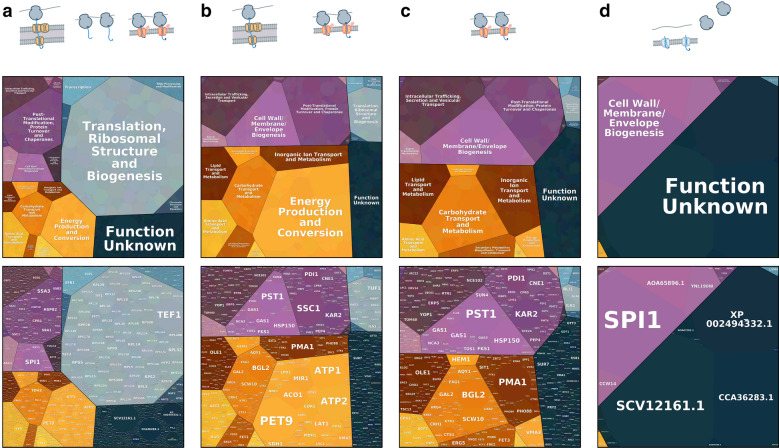
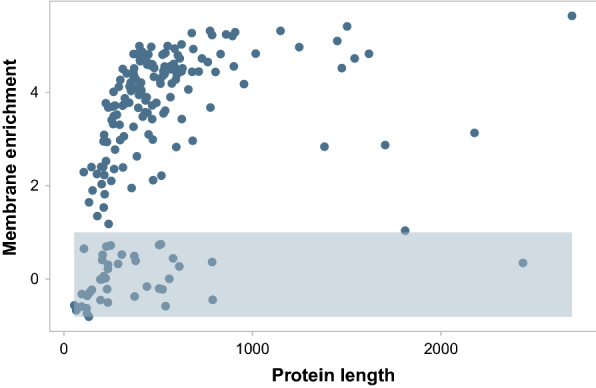
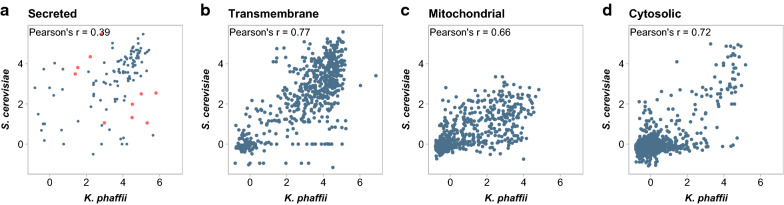
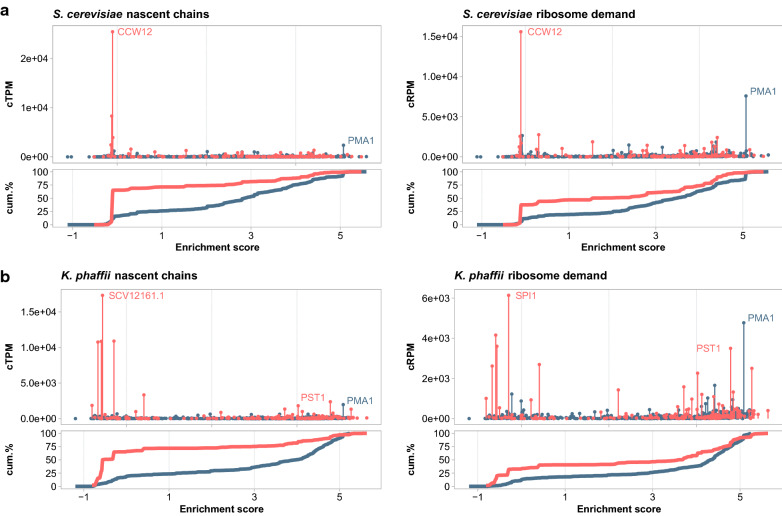
Results: By coupling Ribo-seq with long-read mRNA sequencing, we generated a new annotation of protein-encoding genes. By using Ribo-seq with subcellular fractionation, we quantified demands on co- and posttranslational translocation pathways. During exponential growth in rich media, protein components of the cell-wall represent the greatest number of nascent chains entering the ER. Transcripts encoding the transmembrane protein PMA1 sequester more ribosomes at the ER membrane than any others. Comparison to Saccharomyces cerevisiae reveals conservation in the resources allocated by gene ontology, but variation in the diversity of gene products entering the secretory pathway.
Conclusion: A subset of host proteins, particularly cell-wall components, impose the greatest biosynthetic demands in the early secretory pathway. These proteins are potential targets in strain engineering aimed at alleviating bottlenecks during heterologous protein production.
Keywords: Pichia pastoris; Protein secretion; Resource allocation; Ribosome profiling.
Conflict of interest statement
The authors declare that the current study was funded in part by a gift from Bolt Threads Inc. (Emeryville, CA).
Figures
Similar articles
-
Ribo-seq guided design of enhanced protein secretion in Komagataellaphaffii.Metab Eng. 2025 Sep;91:228-241. doi: 10.1016/j.ymben.2025.04.007. Epub 2025 Apr 30. Metab Eng. 2025. PMID: 40315981
-
Optimization of Multiple Combinations of Early Secretion Pathways to Enhance Extracellular Protein Secretion in Komagataella phaffii.J Agric Food Chem. 2025 Jul 16;73(28):17750-17758. doi: 10.1021/acs.jafc.5c03402. Epub 2025 Jul 5. J Agric Food Chem. 2025. PMID: 40616507
-
Genomic data mining reveals the transaminase repertoire of Komagataella phaffii (Pichia pastoris) strain GS115 and supports a systematic nomenclature.J Genet. 2020;99:49. J Genet. 2020. PMID: 32661202
-
Engineering of the unfolded protein response pathway in Pichia pastoris: enhancing production of secreted recombinant proteins.Appl Microbiol Biotechnol. 2021 Jun;105(11):4397-4414. doi: 10.1007/s00253-021-11336-5. Epub 2021 May 26. Appl Microbiol Biotechnol. 2021. PMID: 34037840 Free PMC article. Review.
-
Ribosomal profiling adds new coding sequences to the proteome.Biochem Soc Trans. 2015 Dec;43(6):1271-6. doi: 10.1042/BST20150170. Biochem Soc Trans. 2015. PMID: 26614672 Review.
Cited by
-
Industrial Production of Proteins with Pichia pastoris-Komagataella phaffii.Biomolecules. 2023 Feb 26;13(3):441. doi: 10.3390/biom13030441. Biomolecules. 2023. PMID: 36979376 Free PMC article. Review.
-
Development of a ribosome profiling protocol to study translation in Kluyveromyces marxianus.FEMS Yeast Res. 2022 Jun 30;22(1):foac024. doi: 10.1093/femsyr/foac024. FEMS Yeast Res. 2022. PMID: 35521744 Free PMC article.
-
CRISPR-Cas9 knockout screen informs efficient reduction of the Komagataella phaffii secretome.Microb Cell Fact. 2024 Jul 31;23(1):217. doi: 10.1186/s12934-024-02466-2. Microb Cell Fact. 2024. PMID: 39085844 Free PMC article.
-
Pathway engineering facilitates efficient protein expression in Pichia pastoris.Appl Microbiol Biotechnol. 2022 Sep;106(18):5893-5912. doi: 10.1007/s00253-022-12139-y. Epub 2022 Aug 30. Appl Microbiol Biotechnol. 2022. PMID: 36040488 Review.
References
MeSH terms
Substances
Supplementary concepts
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
