

Restoring metabolism of myeloid cells reverses cognitive decline in ageing
- PMID: 33473210
- PMCID: PMC8274816
- DOI: 10.1038/s41586-020-03160-0
Restoring metabolism of myeloid cells reverses cognitive decline in ageing
Abstract
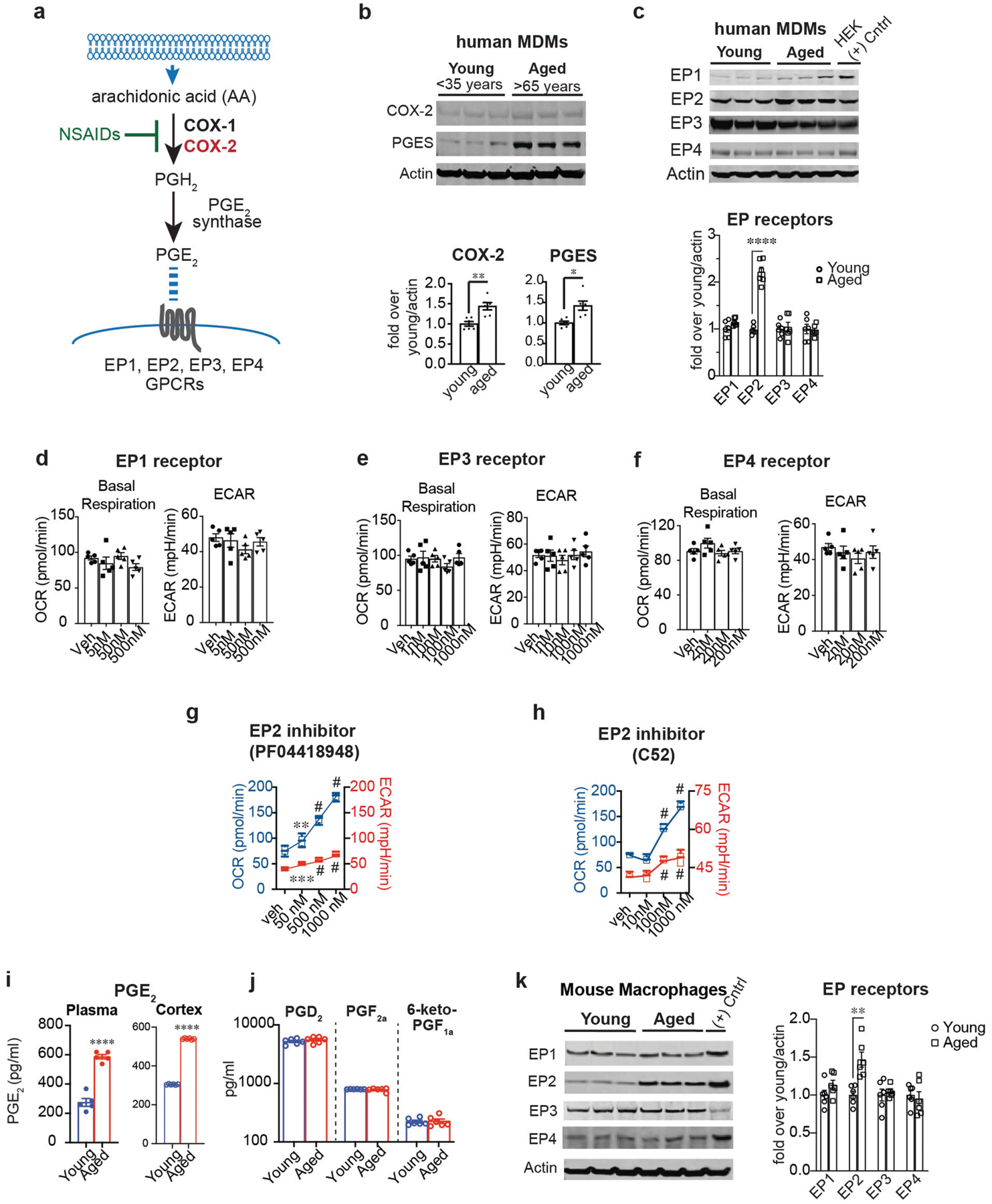
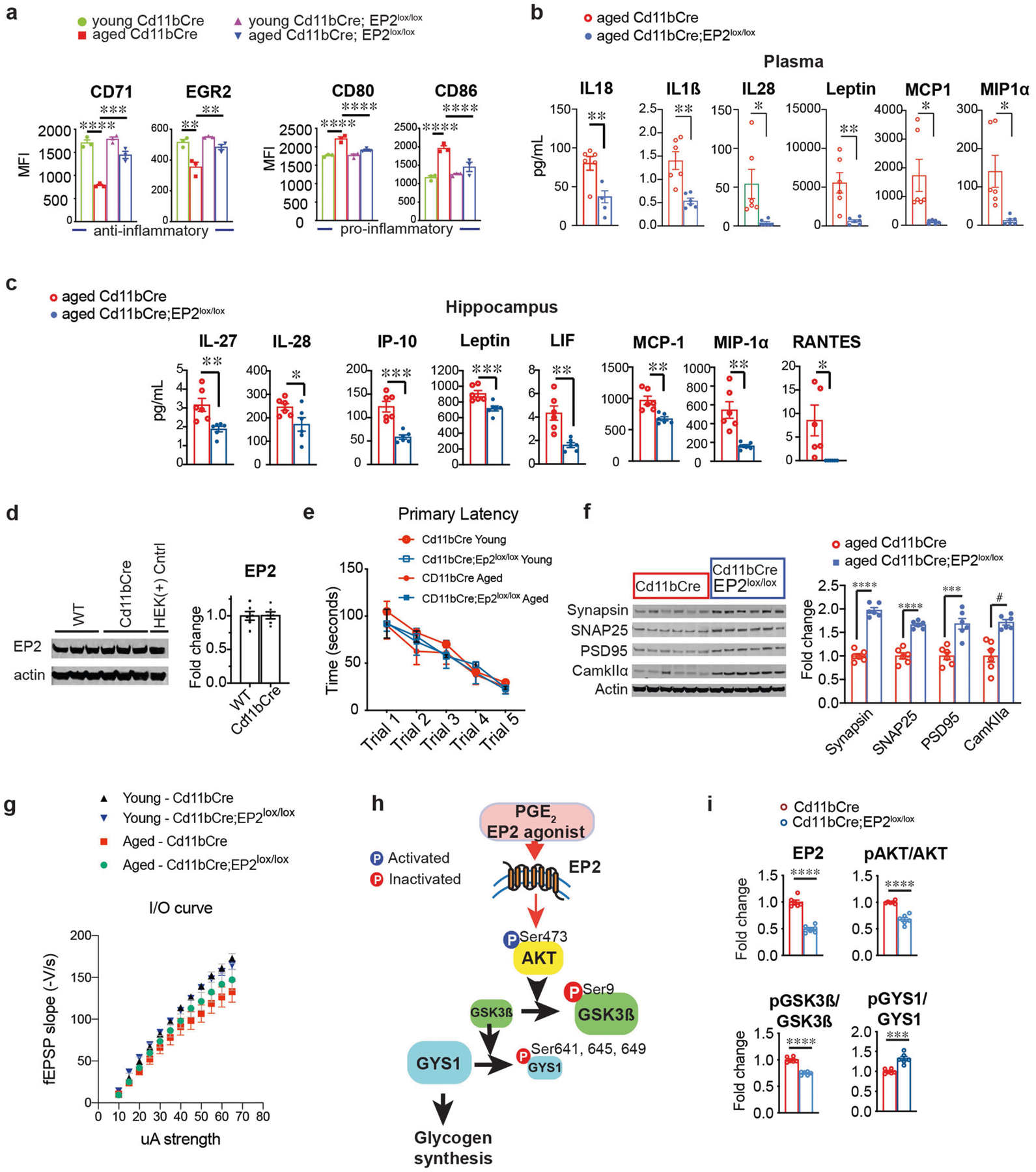
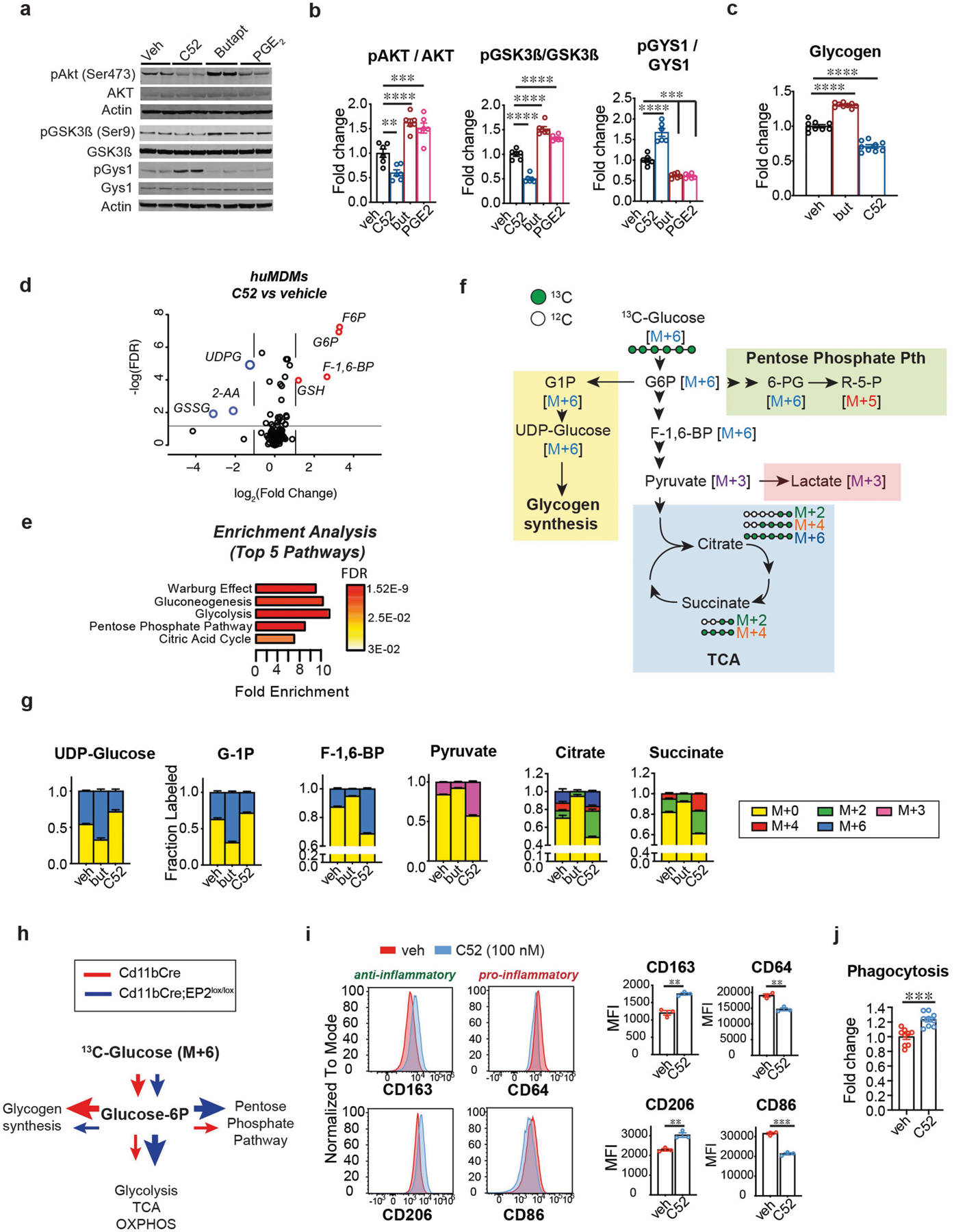
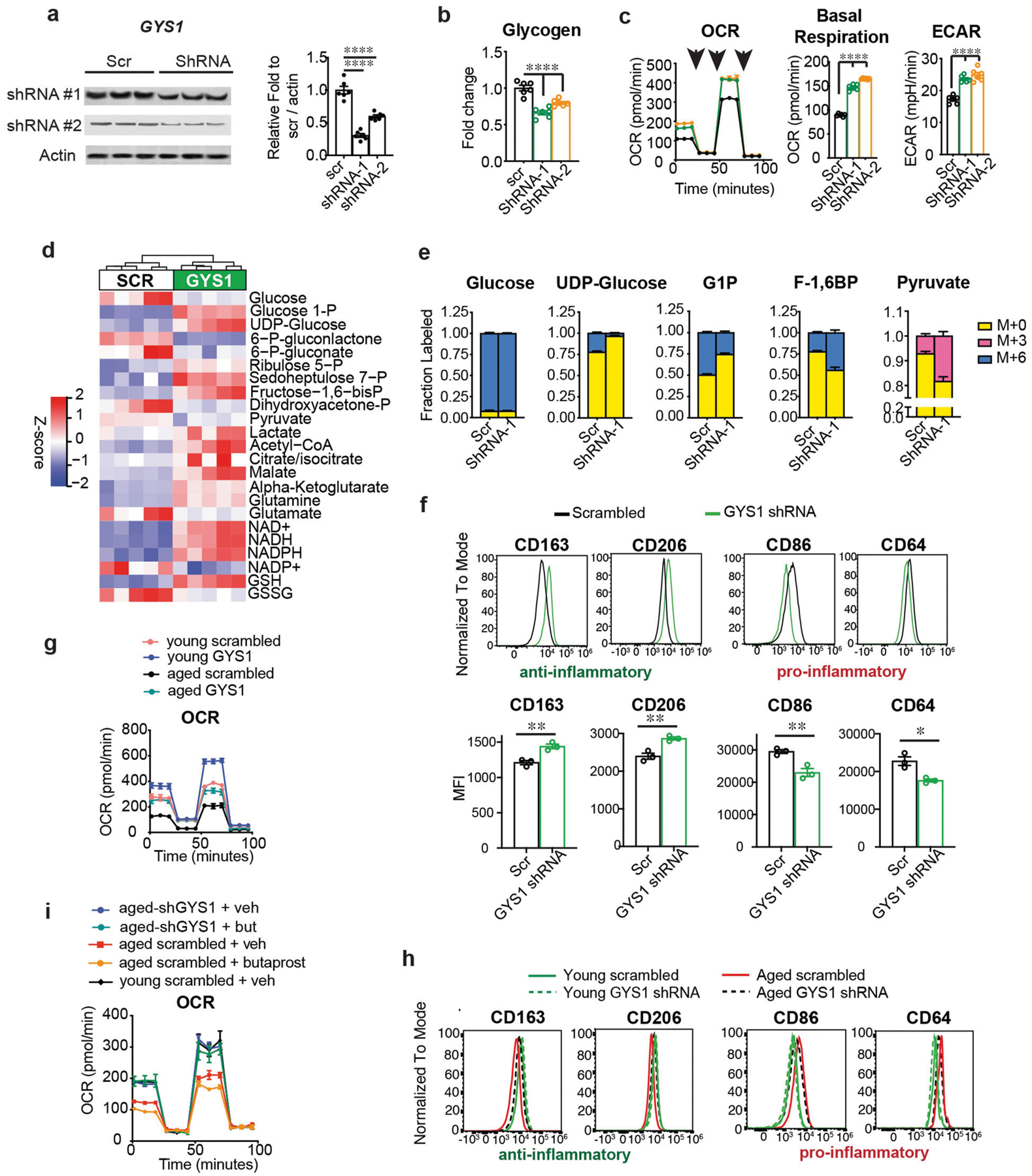
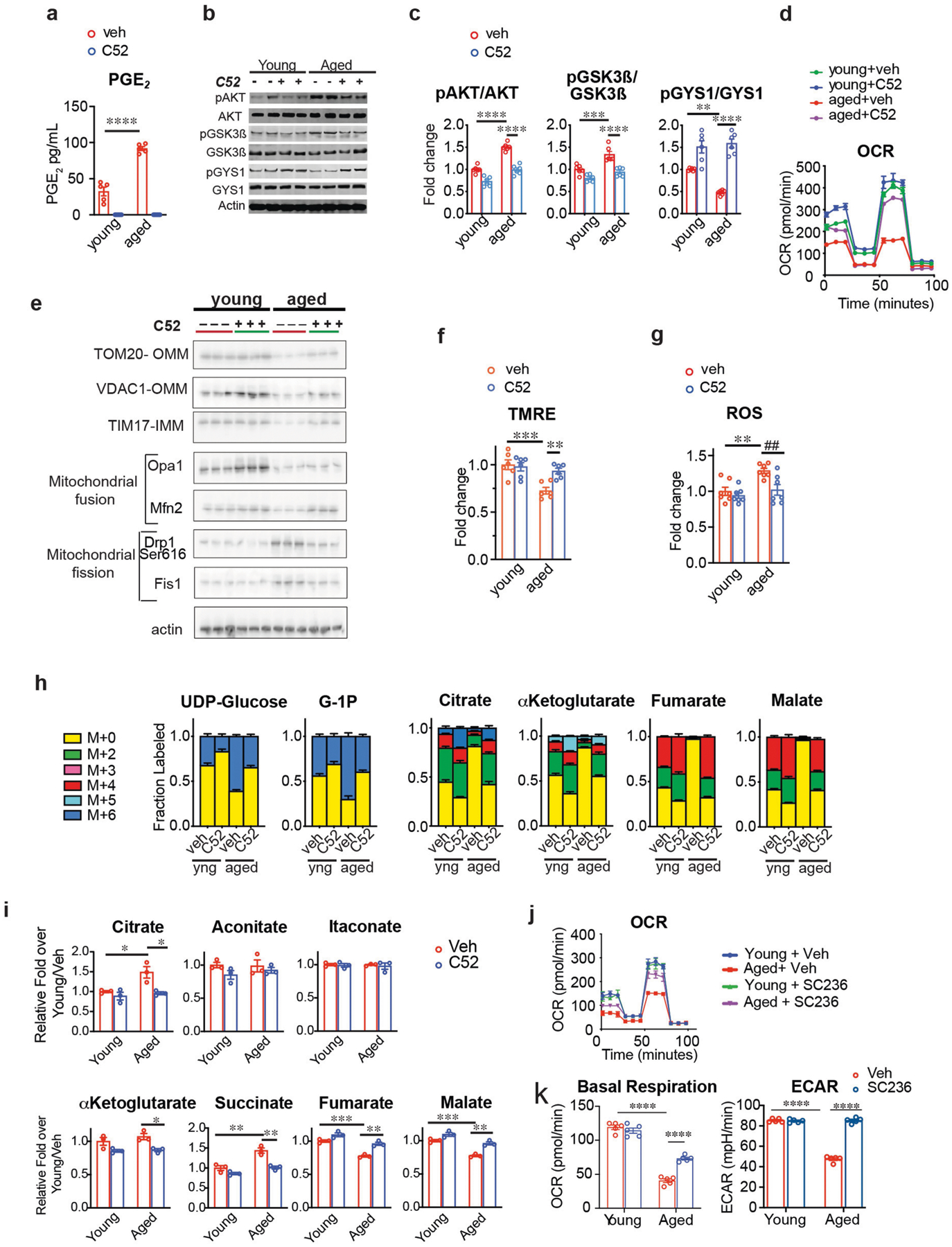
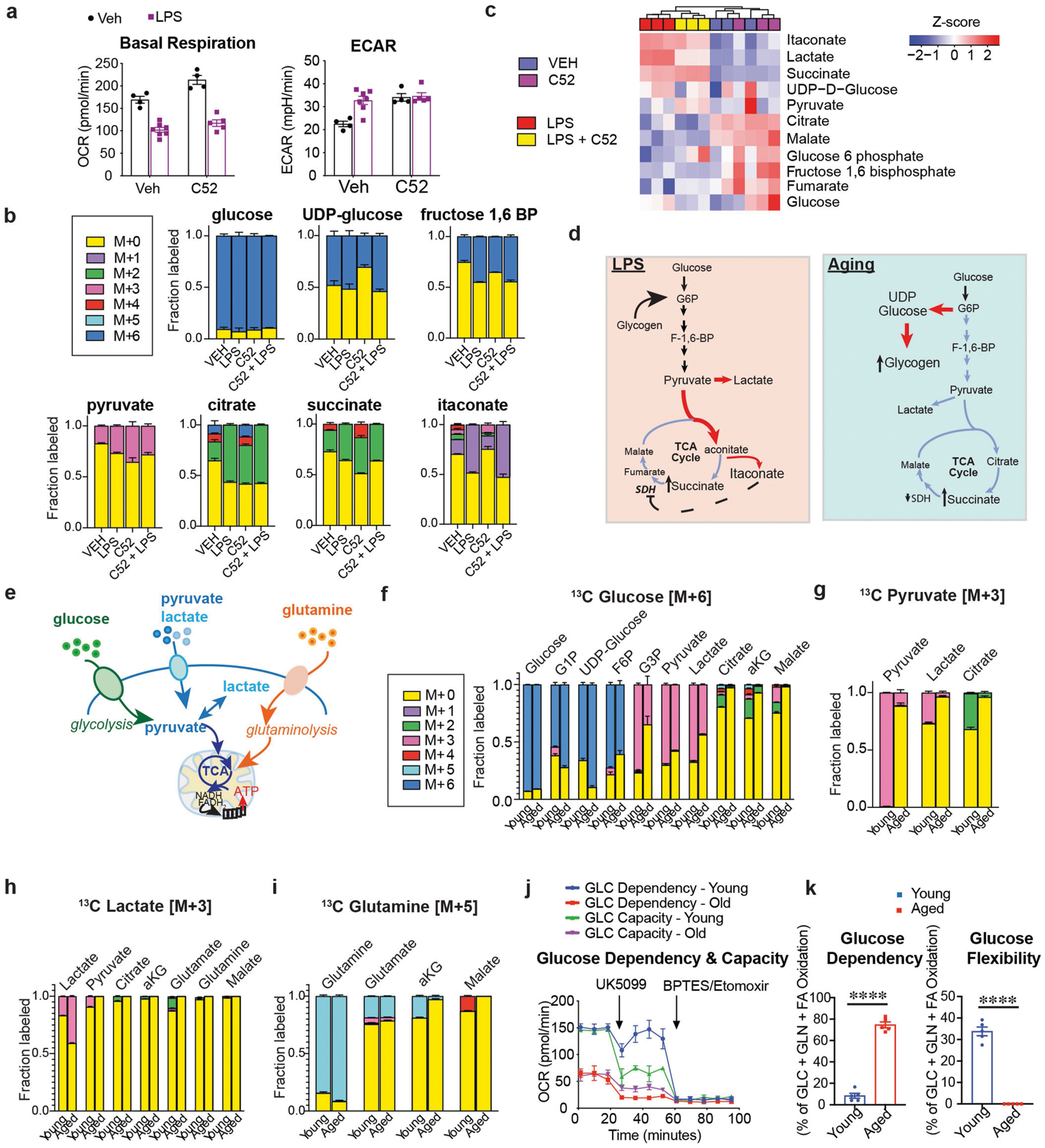
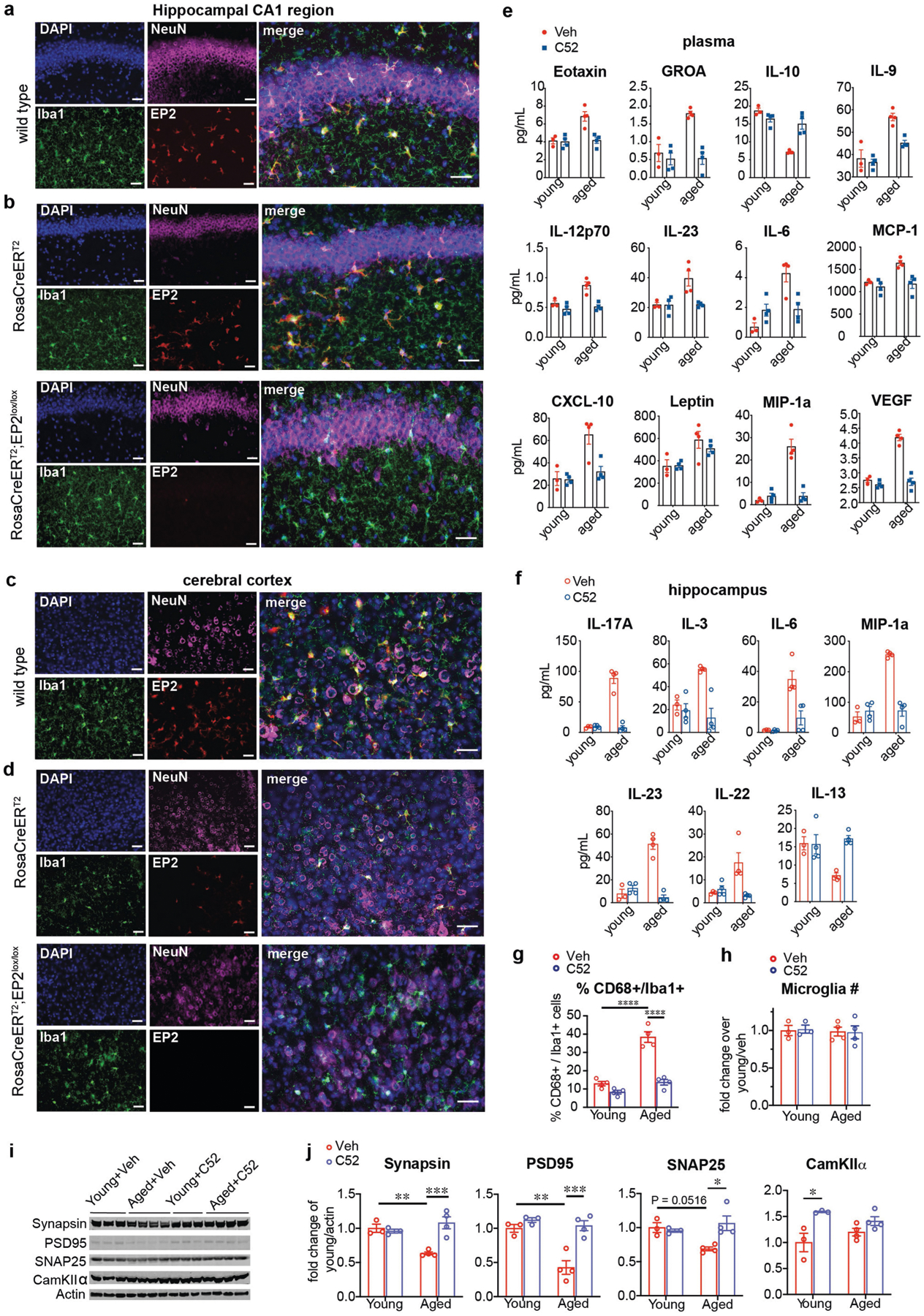
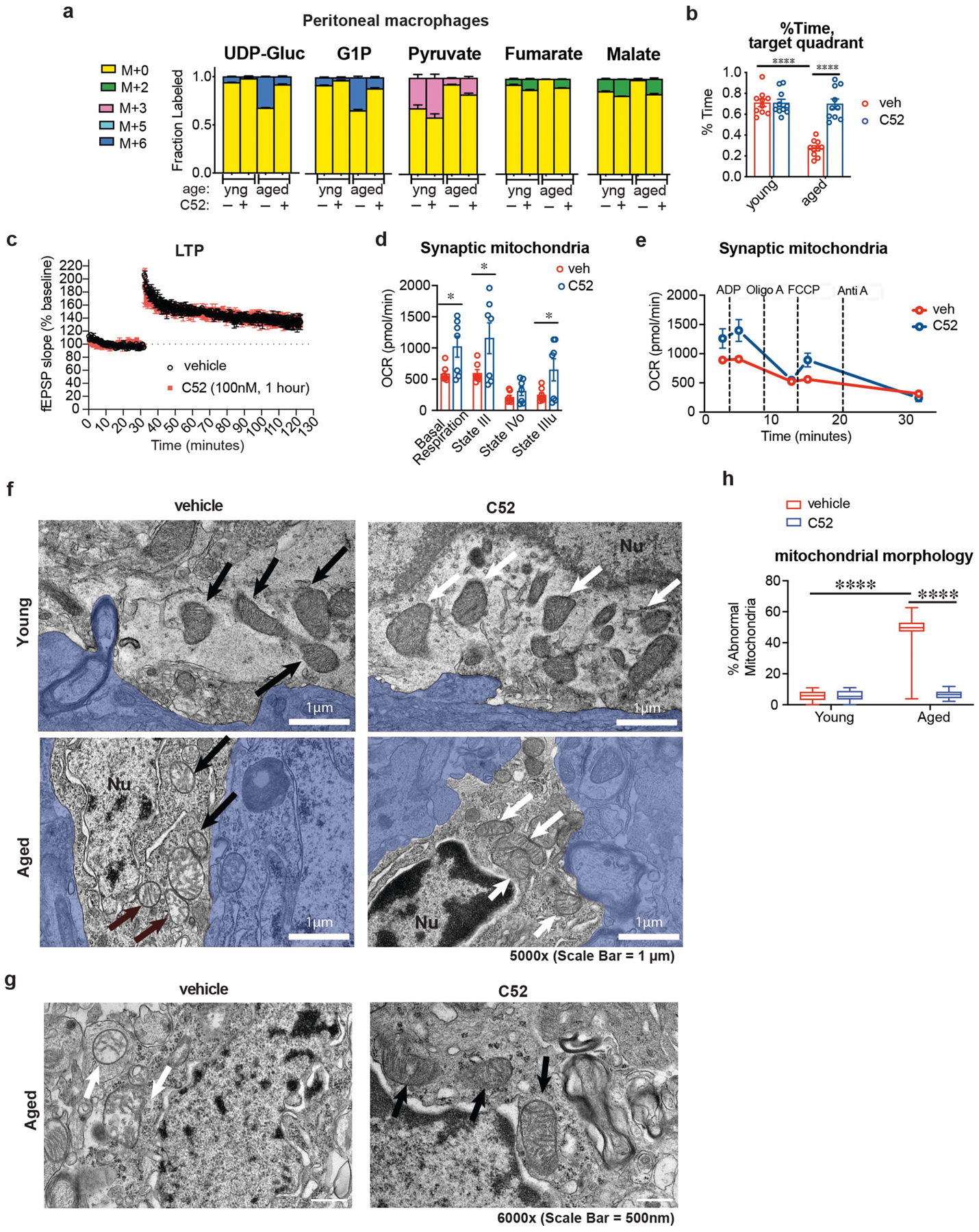
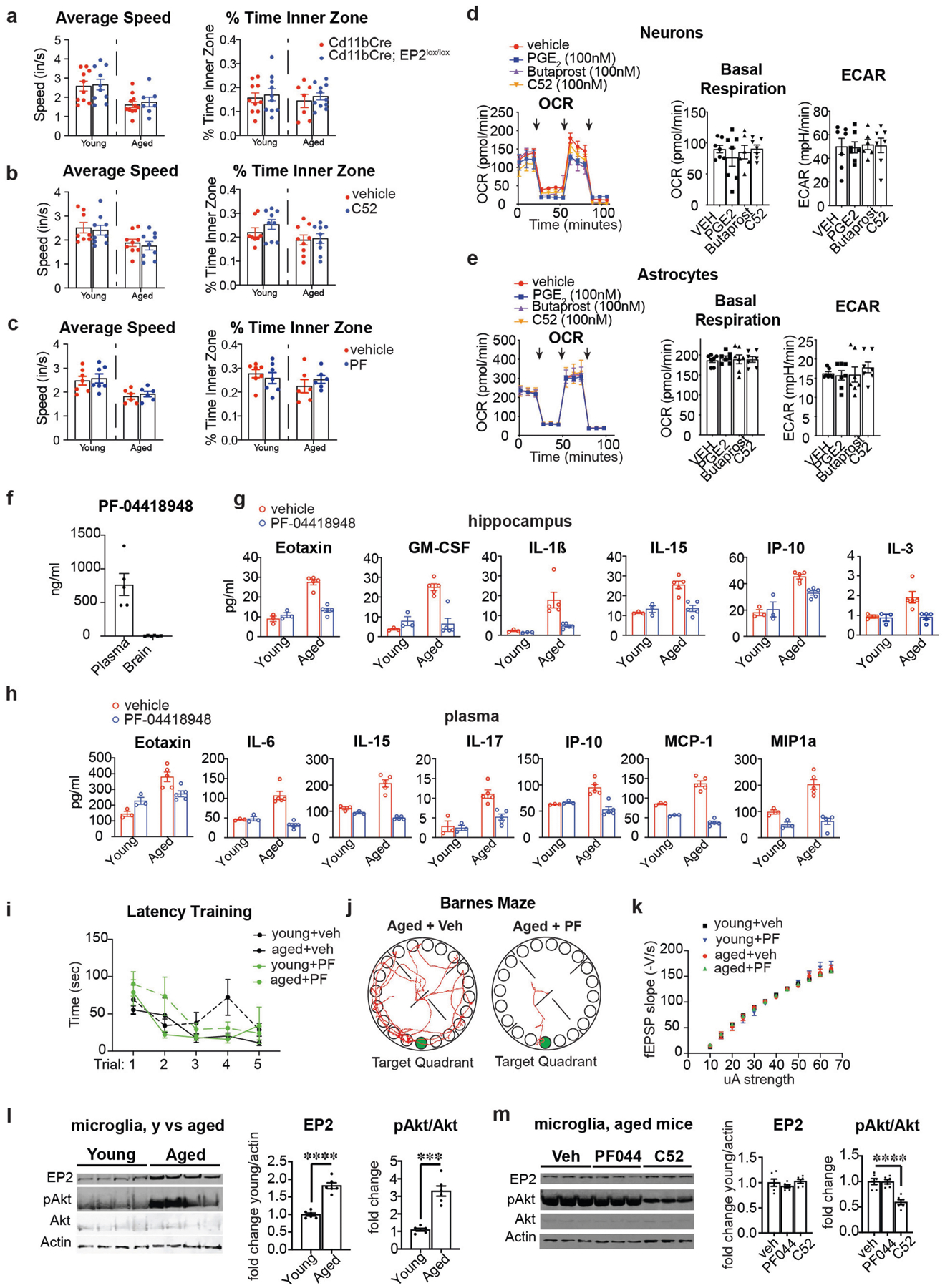
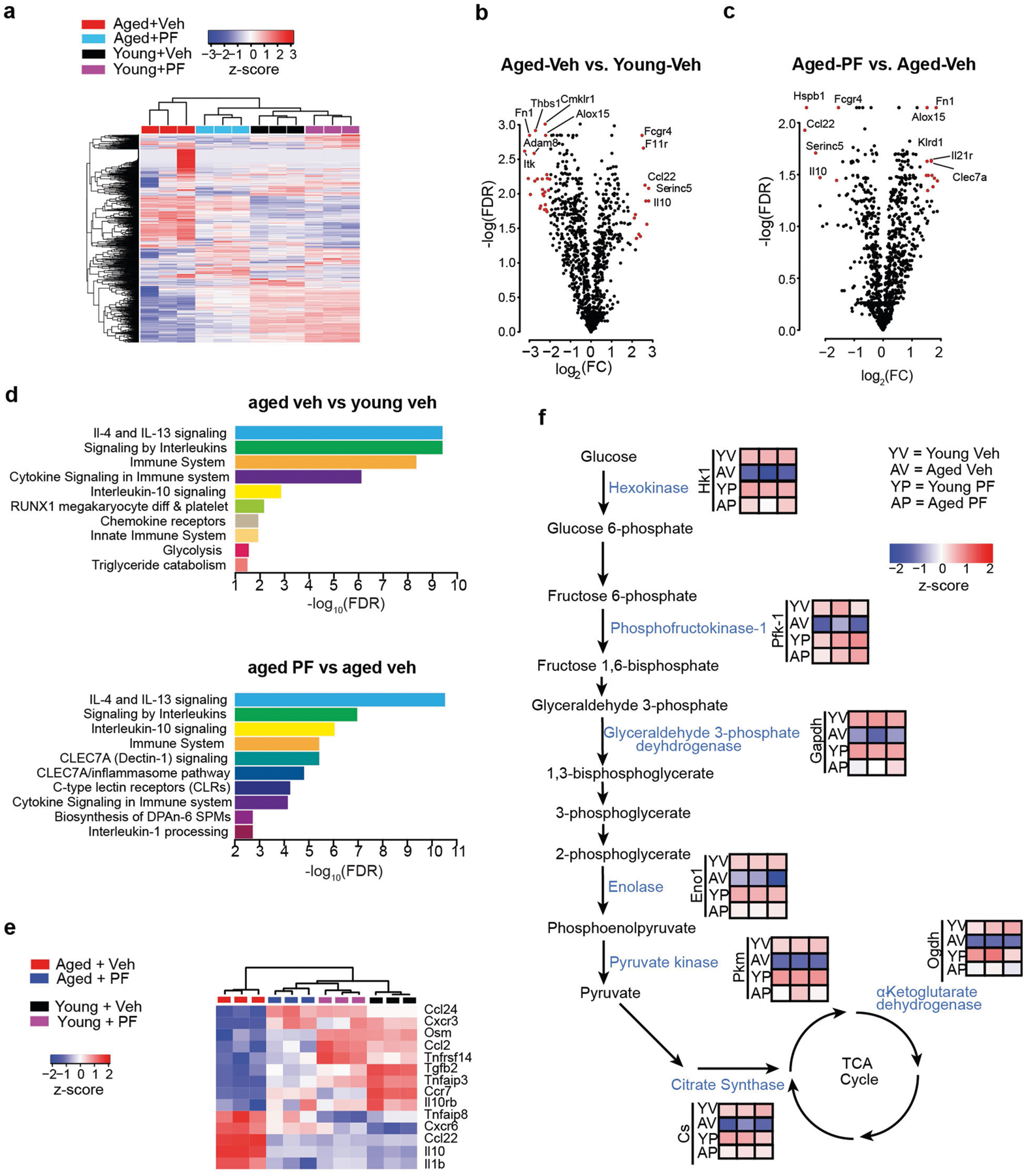
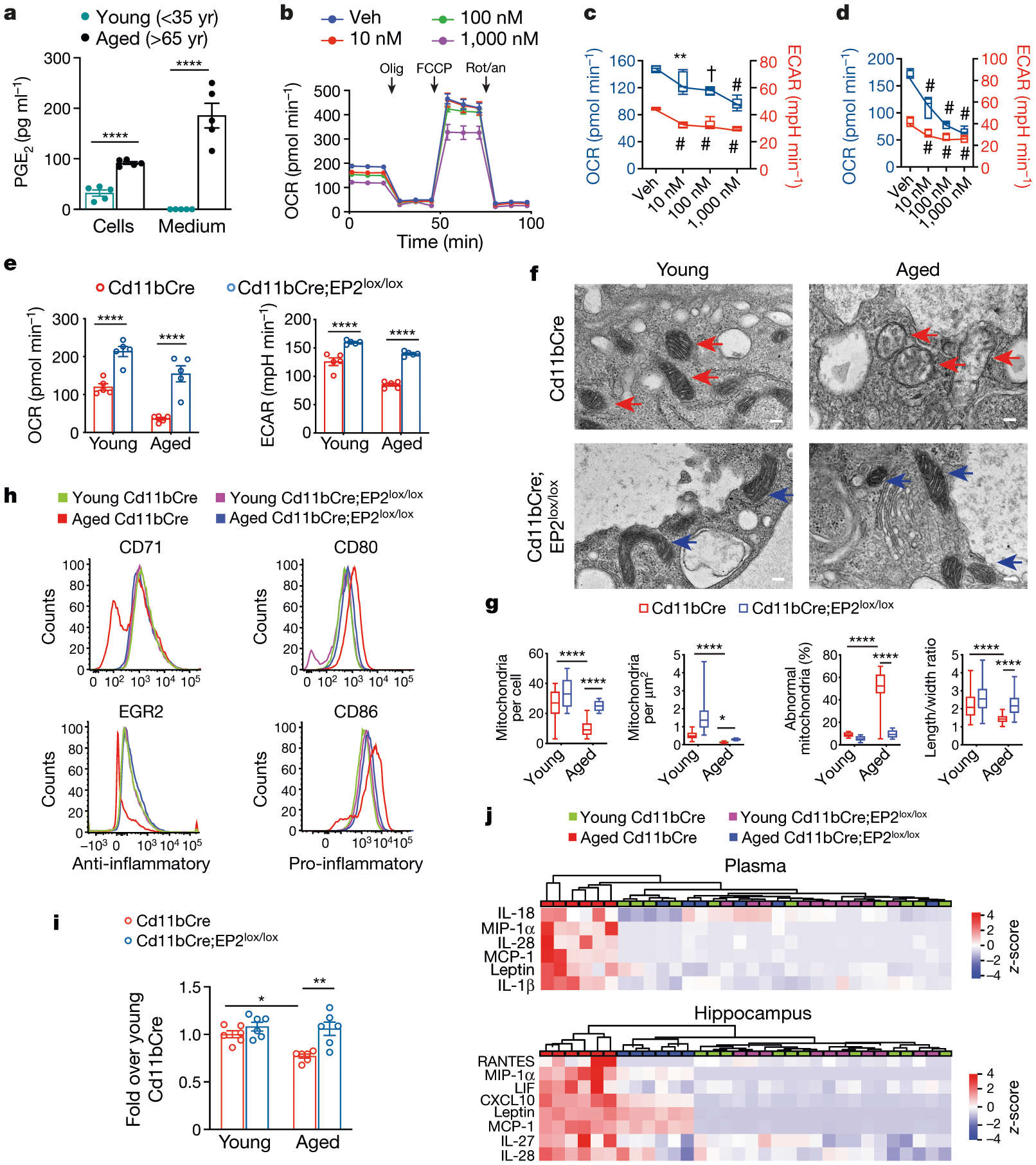
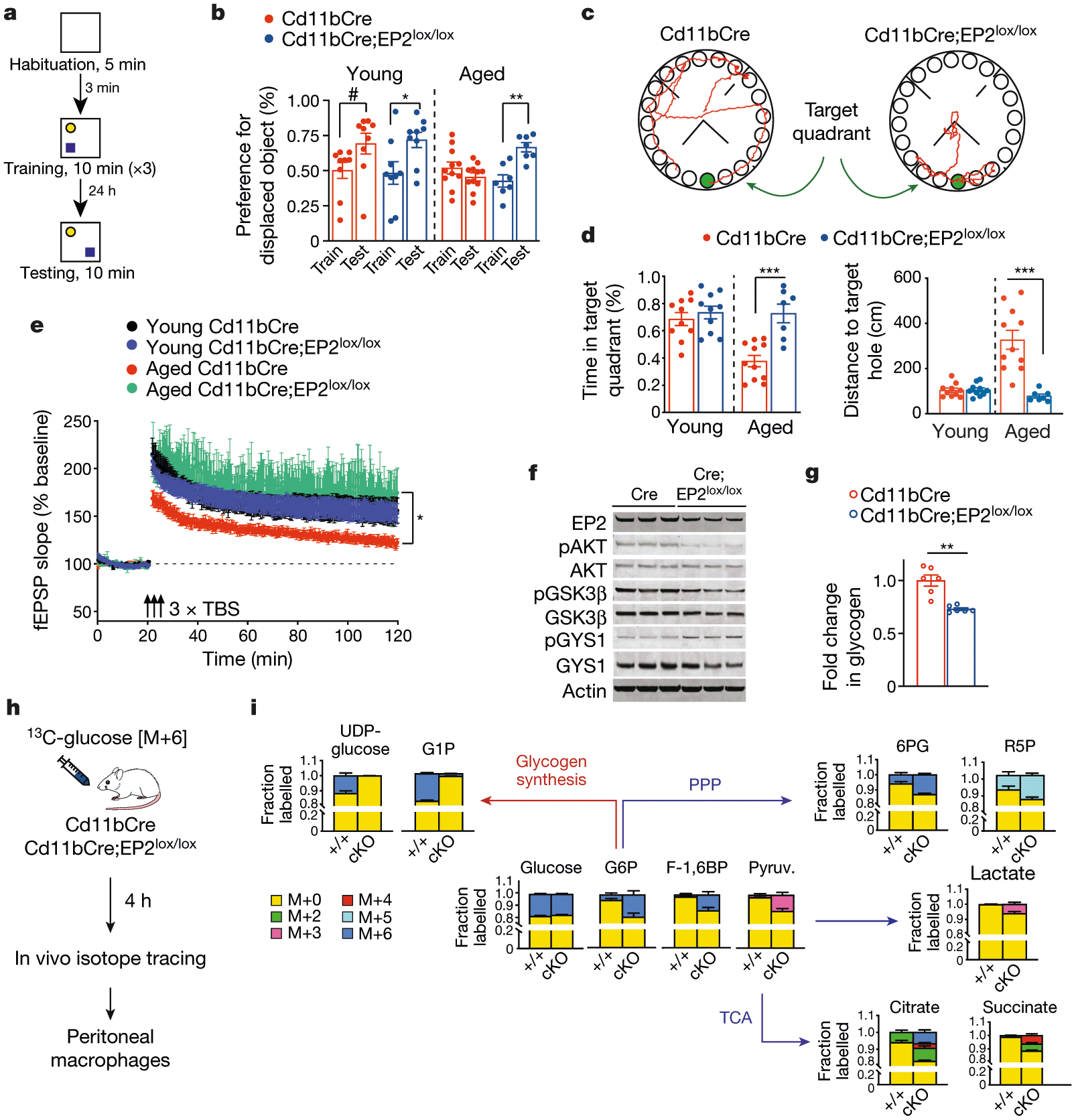
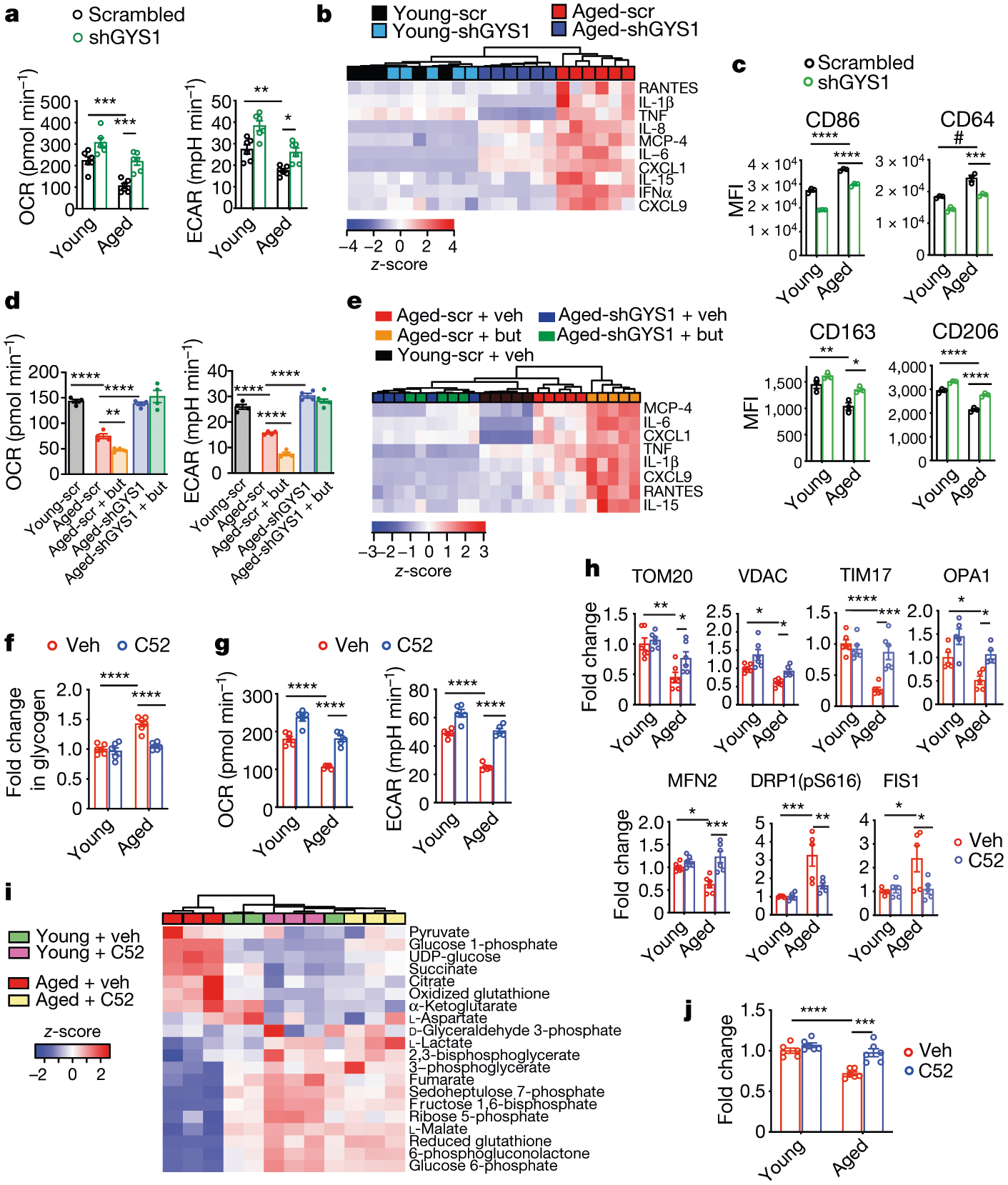
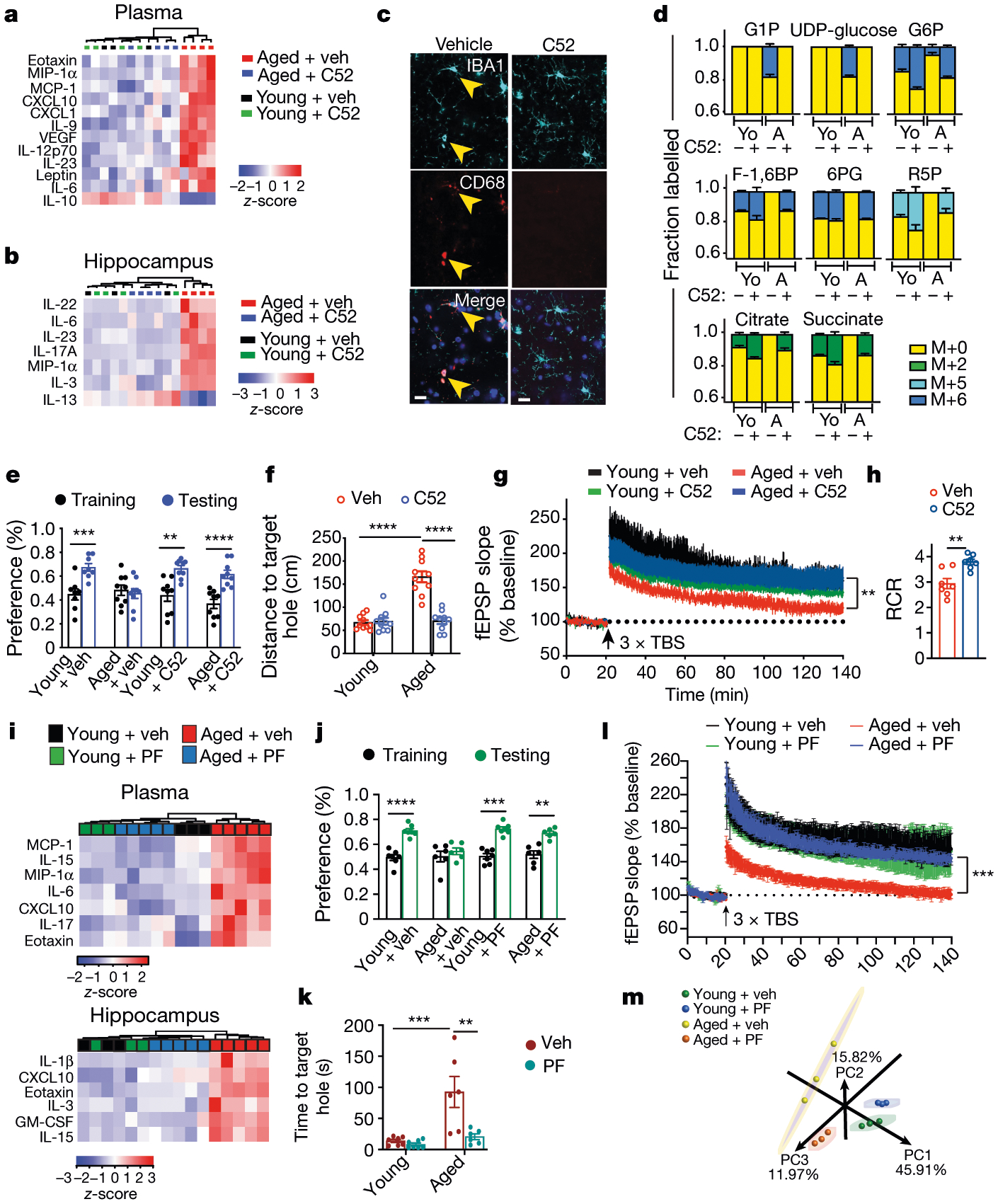
Ageing is characterized by the development of persistent pro-inflammatory responses that contribute to atherosclerosis, metabolic syndrome, cancer and frailty1-3. The ageing brain is also vulnerable to inflammation, as demonstrated by the high prevalence of age-associated cognitive decline and Alzheimer's disease4-6. Systemically, circulating pro-inflammatory factors can promote cognitive decline7,8, and in the brain, microglia lose the ability to clear misfolded proteins that are associated with neurodegeneration9,10. However, the underlying mechanisms that initiate and sustain maladaptive inflammation with ageing are not well defined. Here we show that in ageing mice myeloid cell bioenergetics are suppressed in response to increased signalling by the lipid messenger prostaglandin E2 (PGE2), a major modulator of inflammation11. In ageing macrophages and microglia, PGE2 signalling through its EP2 receptor promotes the sequestration of glucose into glycogen, reducing glucose flux and mitochondrial respiration. This energy-deficient state, which drives maladaptive pro-inflammatory responses, is further augmented by a dependence of aged myeloid cells on glucose as a principal fuel source. In aged mice, inhibition of myeloid EP2 signalling rejuvenates cellular bioenergetics, systemic and brain inflammatory states, hippocampal synaptic plasticity and spatial memory. Moreover, blockade of peripheral myeloid EP2 signalling is sufficient to restore cognition in aged mice. Our study suggests that cognitive ageing is not a static or irrevocable condition but can be reversed by reprogramming myeloid glucose metabolism to restore youthful immune functions.
Conflict of interest statement
Figures
Comment in
-
Reversal of immune-cell shutdown protects the ageing brain.Nature. 2021 Feb;590(7844):44-45. doi: 10.1038/d41586-021-00063-6. Nature. 2021. PMID: 33473186 No abstract available.
-
Teaching old brains new immune tricks.Nat Rev Drug Discov. 2021 Mar;20(3):176. doi: 10.1038/d41573-021-00020-y. Nat Rev Drug Discov. 2021. PMID: 33510472 No abstract available.
-
Teaching old brains new immune tricks.Nat Rev Immunol. 2021 Mar;21(3):133. doi: 10.1038/s41577-021-00515-0. Nat Rev Immunol. 2021. PMID: 33568783 No abstract available.
-
Keep your macrophages fit for healthy aging.Cell Metab. 2021 Mar 2;33(3):468-470. doi: 10.1016/j.cmet.2021.02.004. Cell Metab. 2021. PMID: 33657391
-
Perspectives on Neuroscience and Behavior.Neuroscientist. 2021 Oct;27(5):453. doi: 10.1177/10738584211041575. Neuroscientist. 2021. PMID: 34519550 No abstract available.
References
-
- Franceschi C et al. Inflammaging and anti-inflammaging: a systemic perspective on aging and longevity emerged from studies in humans. Mech. Ageing Dev 128, 92–105 (2007). - PubMed
-
- Nikolich-Žugich J The twilight of immunity: emerging concepts in aging of the immune system. Nat. Immunol 19, 10–19 (2018). - PubMed
-
- Lindenberger U Human cognitive aging: corriger la fortune? Science 346, 572–578 (2014). - PubMed
-
- Brayne C The elephant in the room—healthy brains in later life, epidemiology and public health. Nat. Rev. Neurosci 8, 233–239 (2007). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
