

Resistance to pirimiphos-methyl in West African Anopheles is spreading via duplication and introgression of the Ace1 locus
- PMID: 33476334
- PMCID: PMC7853456
- DOI: 10.1371/journal.pgen.1009253
Resistance to pirimiphos-methyl in West African Anopheles is spreading via duplication and introgression of the Ace1 locus
Abstract
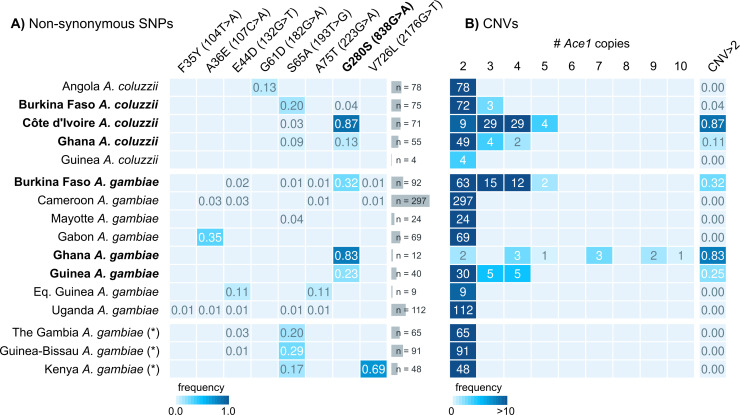
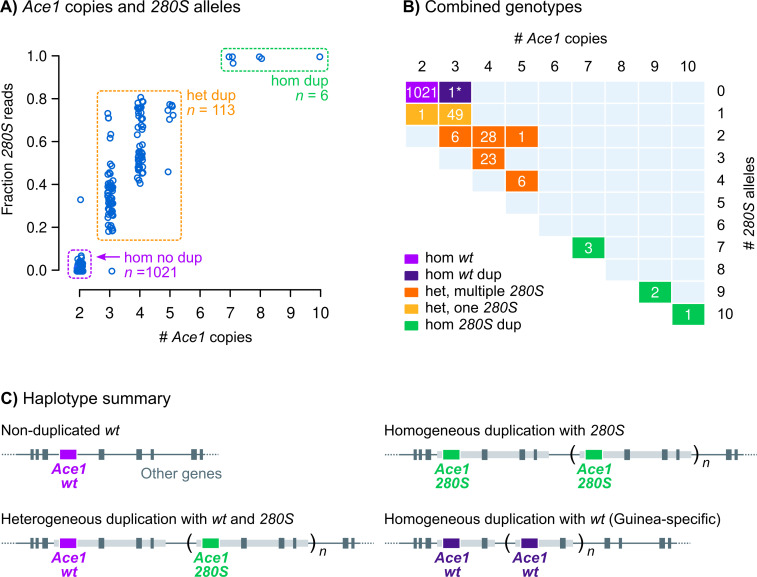
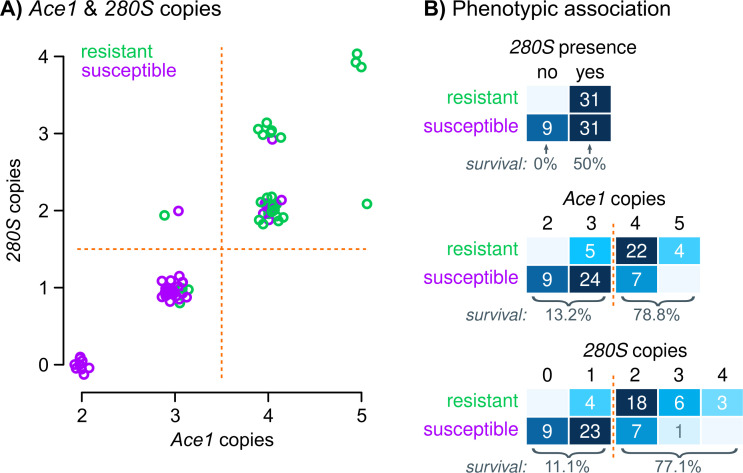
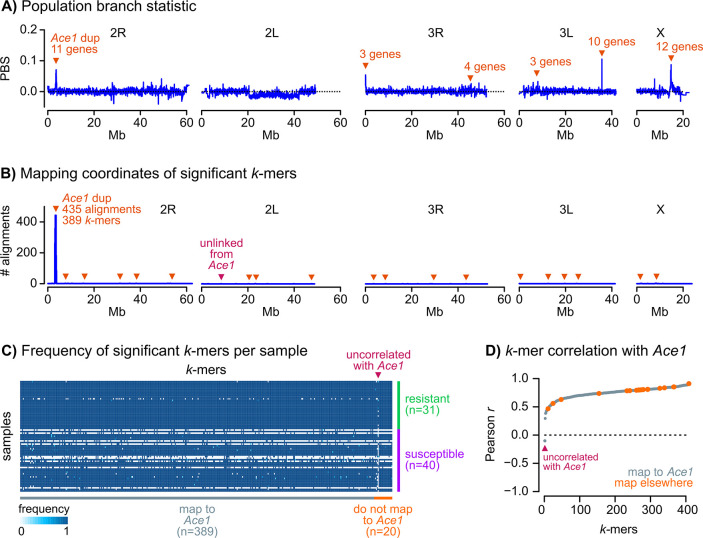
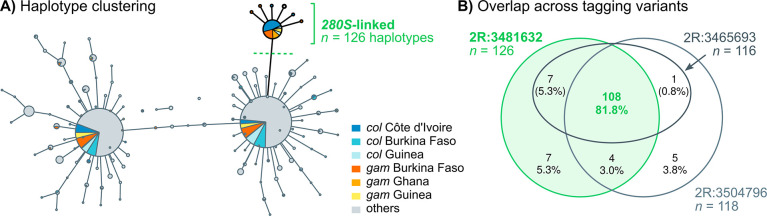
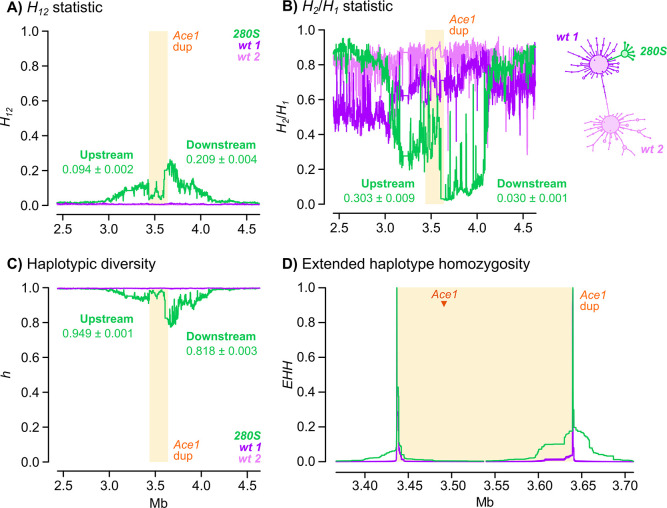
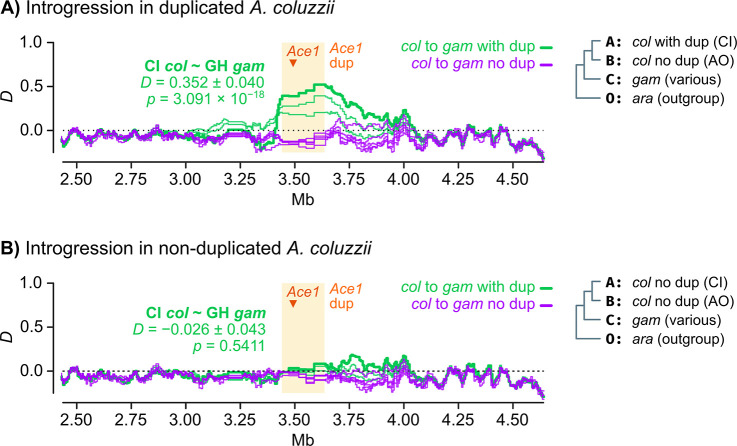
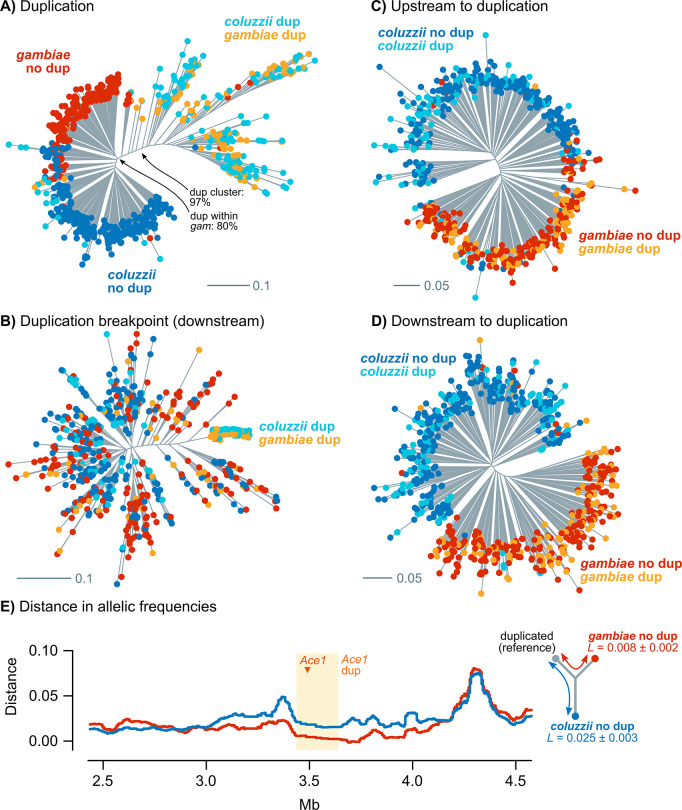
Vector population control using insecticides is a key element of current strategies to prevent malaria transmission in Africa. The introduction of effective insecticides, such as the organophosphate pirimiphos-methyl, is essential to overcome the recurrent emergence of resistance driven by the highly diverse Anopheles genomes. Here, we use a population genomic approach to investigate the basis of pirimiphos-methyl resistance in the major malaria vectors Anopheles gambiae and A. coluzzii. A combination of copy number variation and a single non-synonymous substitution in the acetylcholinesterase gene, Ace1, provides the key resistance diagnostic in an A. coluzzii population from Côte d'Ivoire that we used for sequence-based association mapping, with replication in other West African populations. The Ace1 substitution and duplications occur on a unique resistance haplotype that evolved in A. gambiae and introgressed into A. coluzzii, and is now common in West Africa primarily due to selection imposed by other organophosphate or carbamate insecticides. Our findings highlight the predictive value of this complex resistance haplotype for phenotypic resistance and clarify its evolutionary history, providing tools to for molecular surveillance of the current and future effectiveness of pirimiphos-methyl based interventions.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
References
-
- Dengela D, Seyoum A, Lucas B, Johns B, George K, Belemvire A, et al. Multi-country assessment of residual bio-efficacy of insecticides used for indoor residual spraying in malaria control on different surface types: results from program monitoring in 17 PMI/USAID-supported IRS countries. Parasit Vectors. 2018;11: 71 10.1186/s13071-017-2608-4 - DOI - PMC - PubMed
-
- World Health Organization. Report of the Sixteenth WHOPES Working Group Meeting. Genèva; 2013.
-
- World Health Organization. Global report on insecticide resistance in malaria vectors: 2010–2016. Genèva; 2018.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
