

Vitamin D Modulation of the Innate Immune Response to Paediatric Respiratory Pathogens Associated with Acute Lower Respiratory Infections
- PMID: 33478006
- PMCID: PMC7835957
- DOI: 10.3390/nu13010276
Vitamin D Modulation of the Innate Immune Response to Paediatric Respiratory Pathogens Associated with Acute Lower Respiratory Infections
Abstract
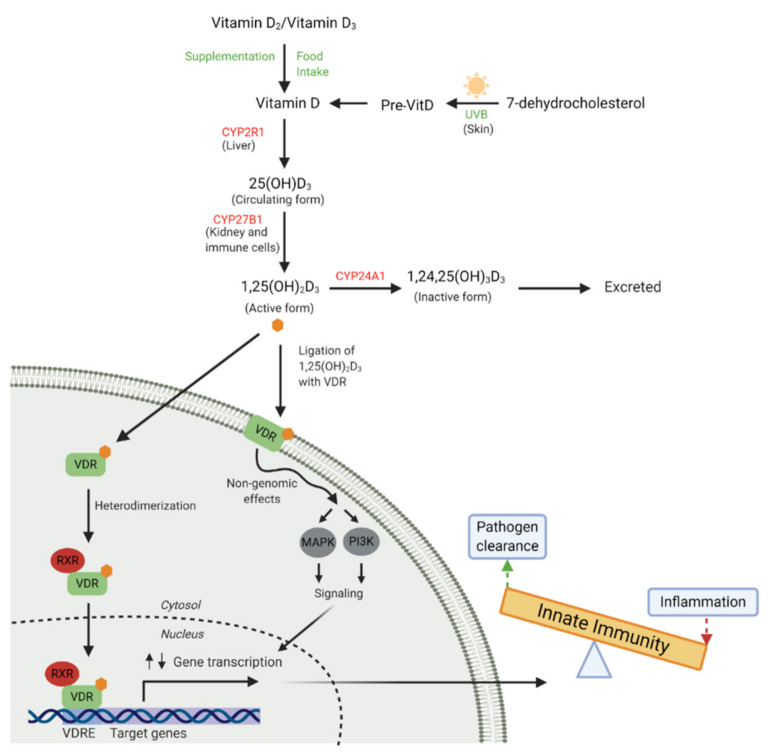
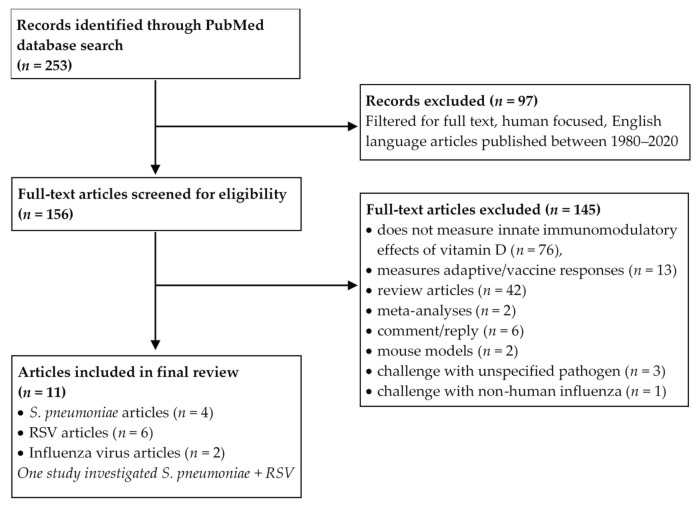
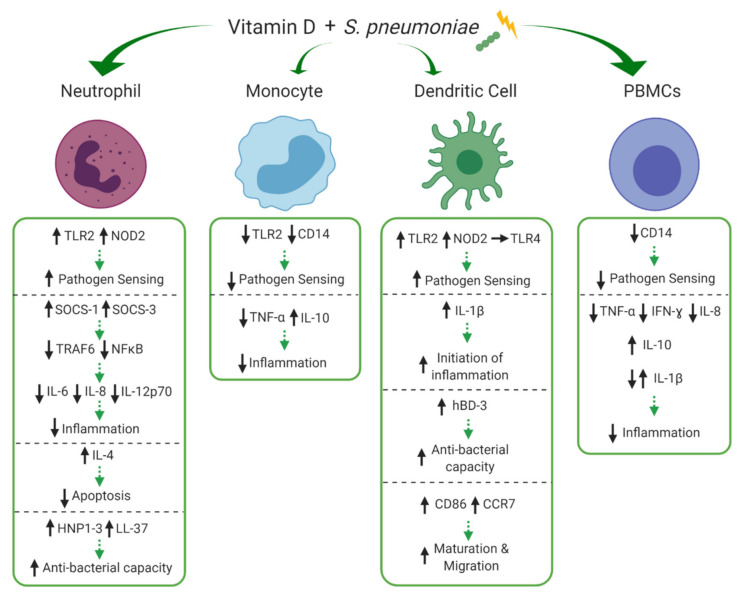
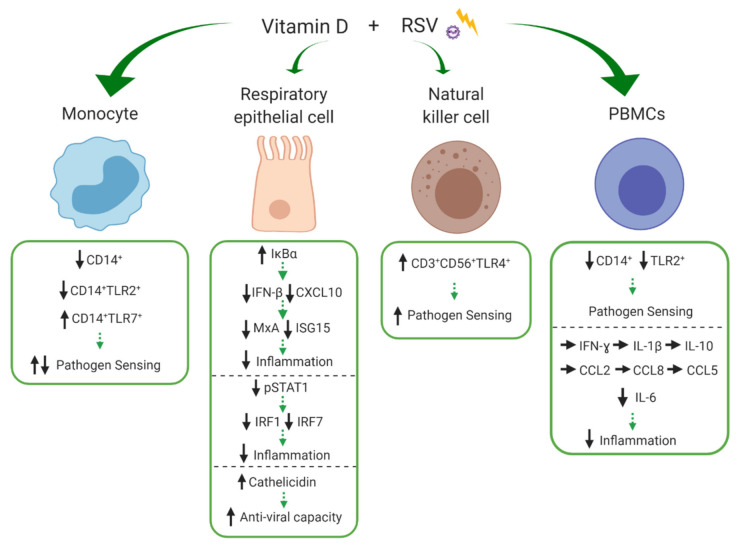
Vitamin D is an essential component of immune function and childhood deficiency is associated with an increased risk of acute lower respiratory infections (ALRIs). Globally, the leading childhood respiratory pathogens are Streptococcus pneumoniae, respiratory syncytial virus and the influenza virus. There is a growing body of evidence describing the innate immunomodulatory properties of vitamin D during challenge with respiratory pathogens, but recent systematic and unbiased synthesis of data is lacking, and future research directions are unclear. We therefore conducted a systematic PubMed literature search using the terms "vitamin D" and "Streptococcus pneumoniae" or "Respiratory Syncytial Virus" or "Influenza". A priori inclusion criteria restricted the review to in vitro studies investigating the effect of vitamin D metabolites on human innate immune cells (primary, differentiated or immortalised) in response to stimulation with the specified respiratory pathogens. Eleven studies met our criteria. Despite some heterogeneity across pathogens and innate cell types, vitamin D modulated pathogen recognition receptor (PRRs: Toll-like receptor 2 (TLR2), TLR4, TLR7 and nucleotide-binding oligomerisation domain-containing protein 2 (NOD2)) expression; increased antimicrobial peptide expression (LL-37, human neutrophil peptide (HNP) 1-3 and β-defensin); modulated autophagosome production reducing apoptosis; and modulated production of inflammatory cytokines (Interleukin (IL) -1β, tumour necrosis factor-α (TNF-α), interferon-ɣ (IFN-ɣ), IL-12p70, IFN-β, Regulated on Activation, Normal T cell Expressed (RANTES), IL-10) and chemokines (IL-8 and C-X-C motif chemokine ligand 10 (CXCL10)). Differential modulation of PRRs and IL-1β was reported across immune cell types; however, this may be due to the experimental design. None of the studies specifically focused on immune responses in cells derived from children. In summary, vitamin D promotes a balanced immune response, potentially enhancing pathogen sensing and clearance and restricting pathogen induced inflammatory dysregulation. This is likely to be important in controlling both ALRIs and the immunopathology associated with poorer outcomes and progression to chronic lung diseases. Many unknowns remain and further investigation is required to clarify the nuances in vitamin D mediated immune responses by pathogen and immune cell type and to determine whether these in vitro findings translate into enhanced immunity and reduced ALRI in the paediatric clinical setting.
Keywords: Streptococcus pneumoniae; acute lower respiratory infections; influenza virus; innate immunity; respiratory syncytial virus; vitamin D.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Naghavi M., Abajobir A.A., Abbafati C., Abbas K.M., Abd-Allah F., Abera S.F., Aboyans V., Adetokunboh O., Afshin A., Agrawal A., et al. Global, regional, and national age-sex specific mortality for 264 causes of death, 1980–2016: A systematic analysis for the Global Burden of Disease Study 2016. Lancet. 2017;390:1151–1210. doi: 10.1016/S0140-6736(17)32152-9. - DOI - PMC - PubMed
-
- Binks M.J., Beissbarth J., Oguoma V.M., Pizzutto S.J., Leach A.J., Smith-Vaughan H.C., McHugh L., Andrews R.M., Webby R., Morris P.S., et al. Acute lower respiratory infections in Indigenous infants in Australia’s Northern Territory across three eras of pneumococcal conjugate vaccine use (2006–15): A population-based cohort study. Lancet Child Adolesc. Health. 2020;4:425–434. doi: 10.1016/S2352-4642(20)30090-0. - DOI - PubMed
-
- A McAllister D., Liu L., Shi T., Chu Y., Reed C., Burrows J., Adeloye D., Rudan I., E Black R., Campbell H., et al. Global, regional, and national estimates of pneumonia morbidity and mortality in children younger than 5 years between 2000 and 2015: A systematic analysis. Lancet Glob. Health. 2019;7:e47–e57. doi: 10.1016/S2214-109X(18)30408-X. - DOI - PMC - PubMed
-
- Histoshi T., McAllister D.A., O’Brien K.L., Simoes E.A.F., Madhi S.A., Gessner B.D., Polack F.P., Balsells E., Acacio S., Aguayo C., et al. Global, regional, and national disease burden estimates of acute lower respiratory infections due to respiratory syncytial virus in young children in 2015: A systematic review and modelling study. Lancet. 2017;390:946–958. doi: 10.1016/s0140-6736(17)30938-8. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Miscellaneous
