

Omega-3 Polyunsaturated Fatty Acids and the Intestinal Epithelium-A Review
- PMID: 33478161
- PMCID: PMC7835870
- DOI: 10.3390/foods10010199
Omega-3 Polyunsaturated Fatty Acids and the Intestinal Epithelium-A Review
Abstract
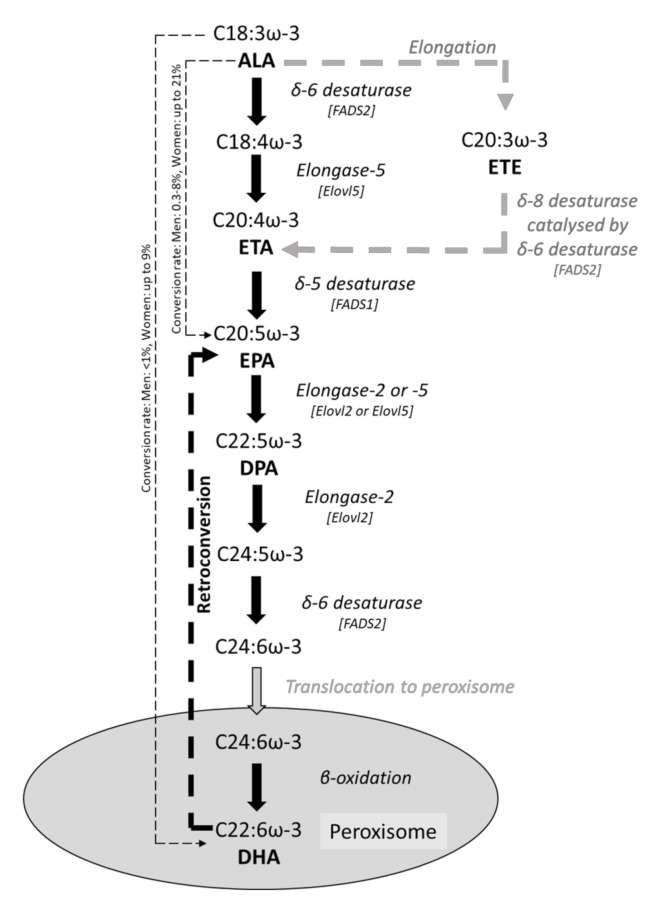
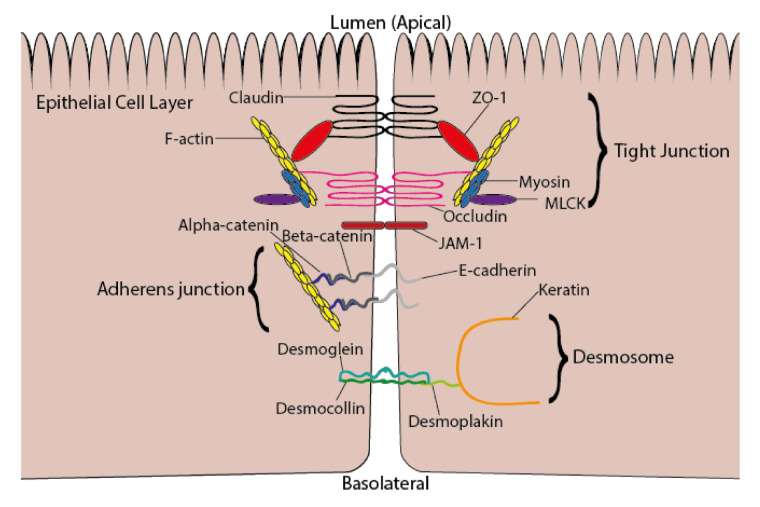
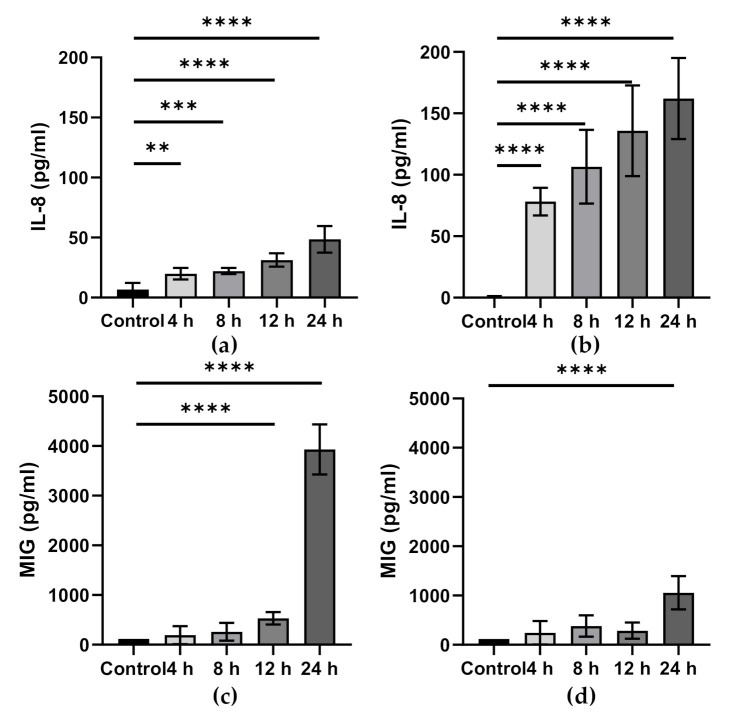
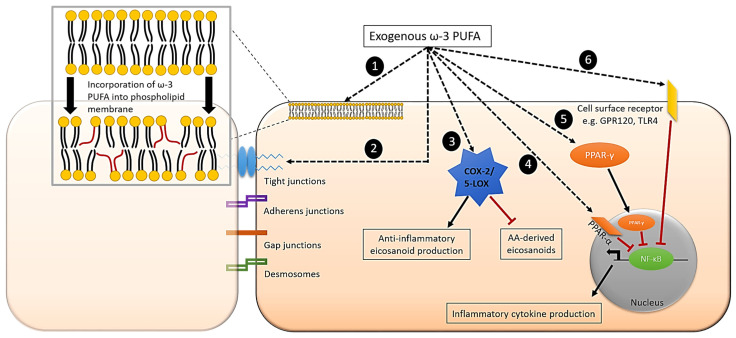
Epithelial cells (enterocytes) form part of the intestinal barrier, the largest human interface between the internal and external environments, and responsible for maintaining regulated intestinal absorption and immunological control. Under inflammatory conditions, the intestinal barrier and its component enterocytes become inflamed, leading to changes in barrier histology, permeability, and chemical mediator production. Omega-3 (ω-3) polyunsaturated fatty acids (PUFAs) can influence the inflammatory state of a range of cell types, including endothelial cells, monocytes, and macrophages. This review aims to assess the current literature detailing the effects of ω-3 PUFAs on epithelial cells. Marine-derived ω-3 PUFAs, eicosapentaenoic acid and docosahexaenoic acid, as well as plant-derived alpha-linolenic acid, are incorporated into intestinal epithelial cell membranes, prevent changes to epithelial permeability, inhibit the production of pro-inflammatory cytokines and eicosanoids and induce the production of anti-inflammatory eicosanoids and docosanoids. Altered inflammatory markers have been attributed to changes in activity and/or expression of proteins involved in inflammatory signalling including nuclear factor kappa-light-chain-enhancer of activated B cells (NF-κB), peroxisome proliferator activated receptor (PPAR) α and γ, G-protein coupled receptor (GPR) 120 and cyclooxygenase (COX)-2. Effective doses for each ω-3 PUFA are difficult to determine due to inconsistencies in dose and time of exposure between different in vitro models and between in vivo and in vitro models. Further research is needed to determine the anti-inflammatory potential of less-studied ω-3 PUFAs, including docosapentaenoic acid and stearidonic acid.
Keywords: chemokine; cytokine; eicosanoid; enterocyte; epithelium; fish oil; inflammation; lipid mediator; permeability; ω-3 PUFA.
Conflict of interest statement
L.A.D. and C.E.C. declare no conflict of interest. P.C.C. acts as a consultant to BASF AS, Smartfish, DSM, Cargill, Danone/Nutricia and Fresenius-Kabi.
Figures
References
Publication types
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
