

Glycolysis fuels phosphoinositide 3-kinase signaling to bolster T cell immunity
- PMID: 33479154
- PMCID: PMC8380312
- DOI: 10.1126/science.abb2683
Glycolysis fuels phosphoinositide 3-kinase signaling to bolster T cell immunity
Abstract
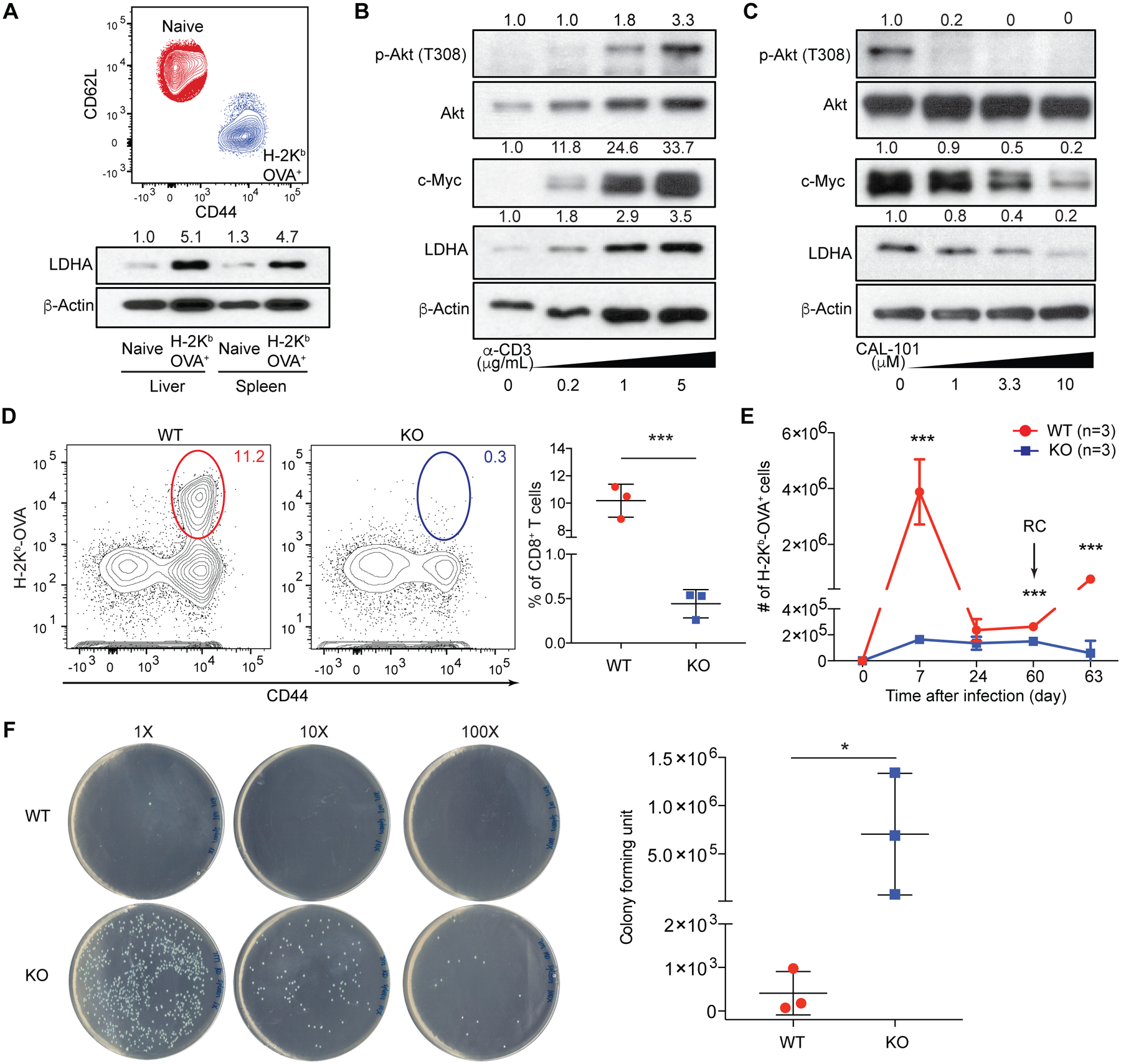
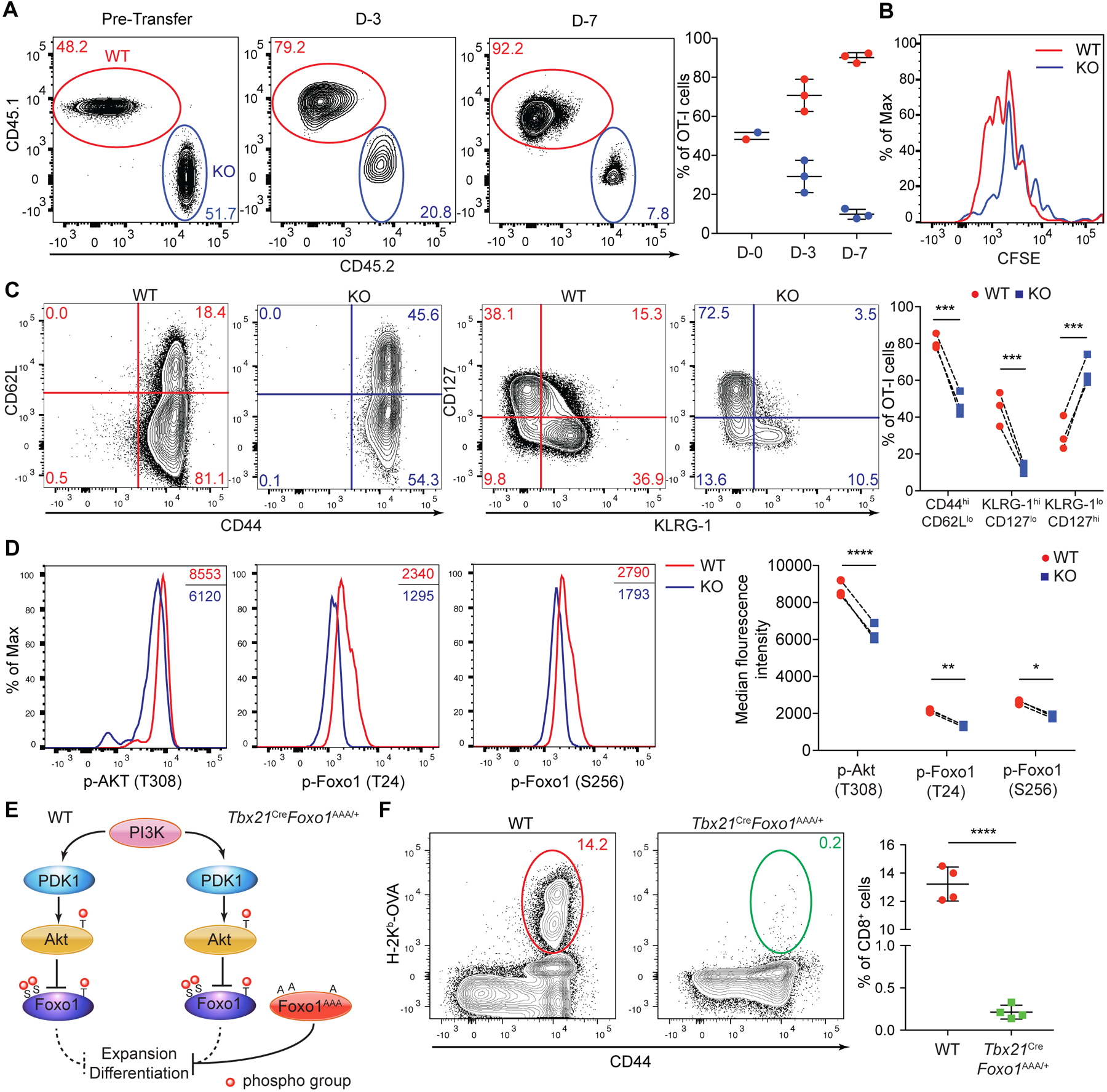
Infection triggers expansion and effector differentiation of T cells specific for microbial antigens in association with metabolic reprograming. We found that the glycolytic enzyme lactate dehydrogenase A (LDHA) is induced in CD8+ T effector cells through phosphoinositide 3-kinase (PI3K) signaling. In turn, ablation of LDHA inhibits PI3K-dependent phosphorylation of Akt and its transcription factor target Foxo1, causing defective antimicrobial immunity. LDHA deficiency cripples cellular redox control and diminishes adenosine triphosphate (ATP) production in effector T cells, resulting in attenuated PI3K signaling. Thus, nutrient metabolism and growth factor signaling are highly integrated processes, with glycolytic ATP serving as a rheostat to gauge PI3K-Akt-Foxo1 signaling in the control of T cell immunity. Such a bioenergetic mechanism for the regulation of signaling may explain the Warburg effect.
Copyright © 2021, American Association for the Advancement of Science.
Conflict of interest statement
Figures


References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
