

White pupae phenotype of tephritids is caused by parallel mutations of a MFS transporter
- PMID: 33479218
- PMCID: PMC7820335
- DOI: 10.1038/s41467-020-20680-5
White pupae phenotype of tephritids is caused by parallel mutations of a MFS transporter
Abstract
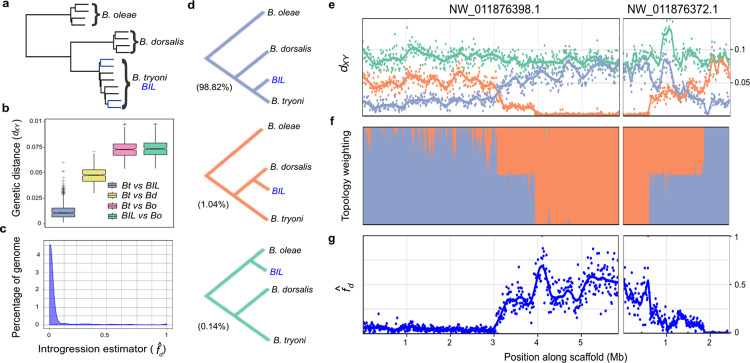
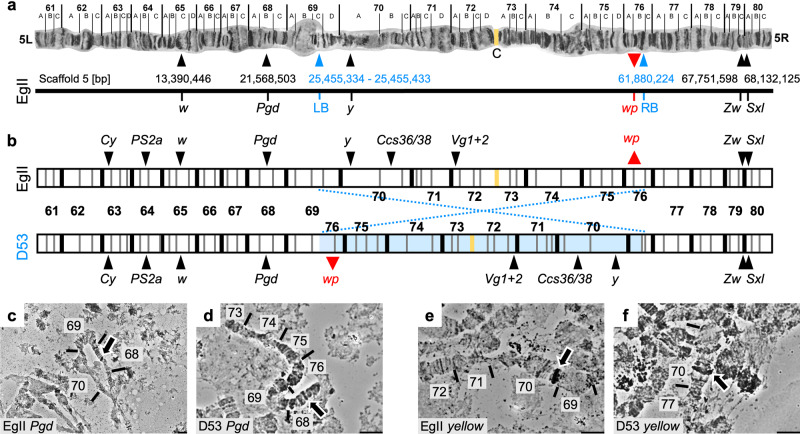
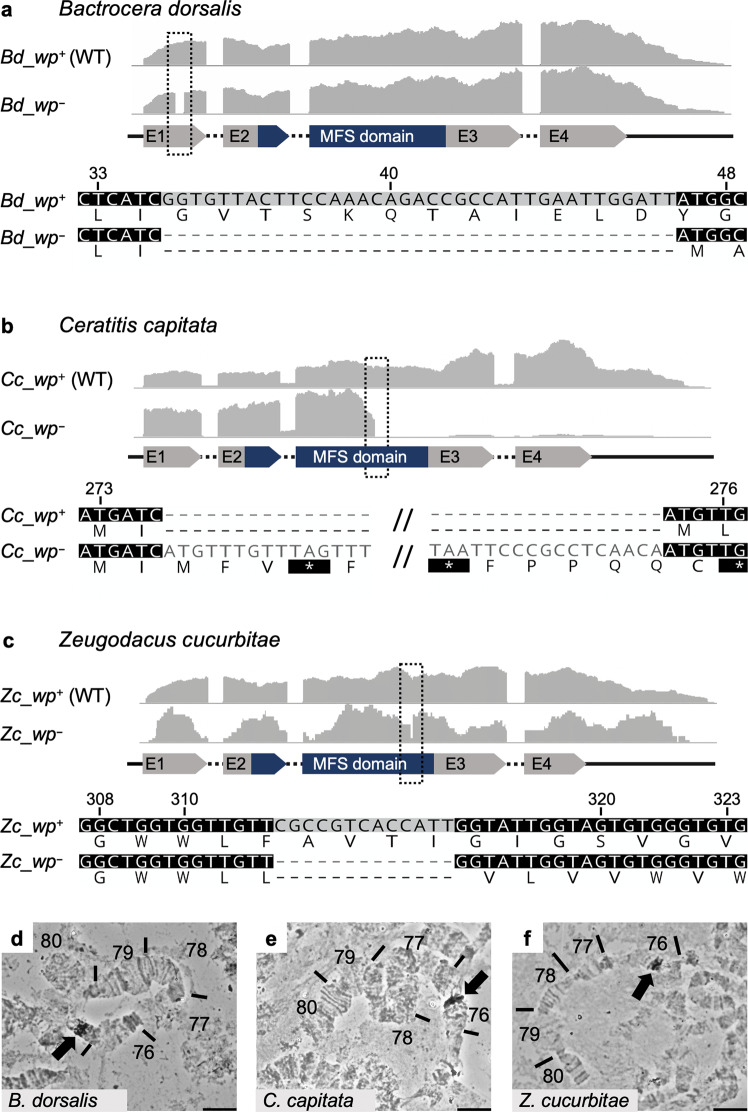
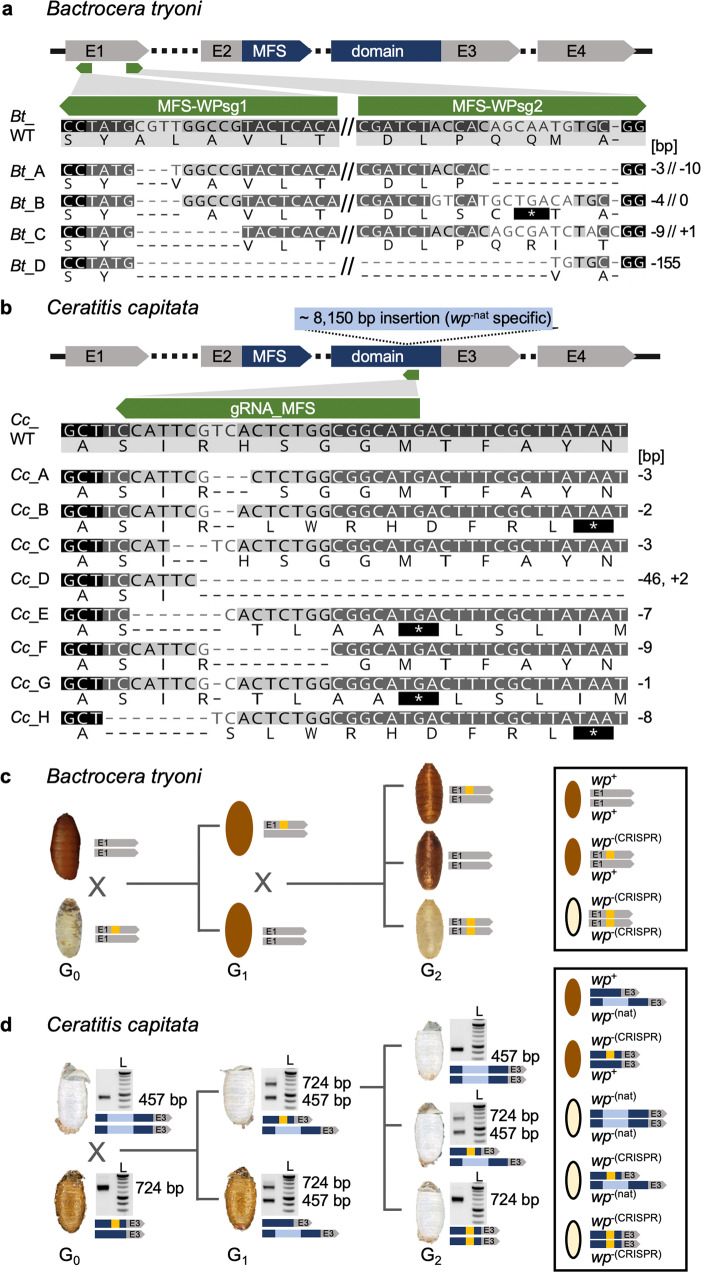
Mass releases of sterilized male insects, in the frame of sterile insect technique programs, have helped suppress insect pest populations since the 1950s. In the major horticultural pests Bactrocera dorsalis, Ceratitis capitata, and Zeugodacus cucurbitae, a key phenotype white pupae (wp) has been used for decades to selectively remove females before releases, yet the gene responsible remained unknown. Here, we use classical and modern genetic approaches to identify and functionally characterize causal wp- mutations in these distantly related fruit fly species. We find that the wp phenotype is produced by parallel mutations in a single, conserved gene. CRISPR/Cas9-mediated knockout of the wp gene leads to the rapid generation of white pupae strains in C. capitata and B. tryoni. The conserved phenotype and independent nature of wp- mutations suggest this technique can provide a generic approach to produce sexing strains in other major medical and agricultural insect pests.
Conflict of interest statement
The authors declare no competing interests.
Figures
Similar articles
-
Deep orange gene editing triggers temperature-sensitive lethal phenotypes in Ceratitis capitata.BMC Biotechnol. 2024 Feb 1;24(1):7. doi: 10.1186/s12896-024-00832-x. BMC Biotechnol. 2024. PMID: 38302991 Free PMC article.
-
Highly efficient genome editing by homology-directed repair using Cas9 protein in Ceratitis capitata.Insect Biochem Mol Biol. 2018 Oct;101:85-93. doi: 10.1016/j.ibmb.2018.08.004. Epub 2018 Aug 26. Insect Biochem Mol Biol. 2018. PMID: 30157456
-
Improving the Phenotypic Properties of the Ceratitis capitata (Diptera: Tephritidae) Temperature-Sensitive Lethal Genetic Sexing Strain in Support of Sterile Insect Technique Applications.J Econ Entomol. 2020 Dec 9;113(6):2688-2694. doi: 10.1093/jee/toaa220. J Econ Entomol. 2020. PMID: 33020821 Free PMC article.
-
Genetic sexing strains in medfly, Ceratitis capitata, sterile insect technique programmes.Genetica. 2002 Sep;116(1):5-13. doi: 10.1023/a:1020951407069. Genetica. 2002. PMID: 12484522 Review.
-
How functional genomics will impact fruit fly pest control: the example of the Mediterranean fruit fly, Ceratitis capitata.BMC Genet. 2014;15 Suppl 2(Suppl 2):S11. doi: 10.1186/1471-2156-15-S2-S11. Epub 2014 Dec 1. BMC Genet. 2014. PMID: 25471105 Free PMC article. Review.
Cited by
-
Development of a novel genetic sexing strain of Ceratitis capitata based on an X-autosome translocation.Sci Rep. 2023 Sep 27;13(1):16167. doi: 10.1038/s41598-023-43164-0. Sci Rep. 2023. PMID: 37758733 Free PMC article.
-
Genomic and cytogenetic analysis of the Ceratitis capitata temperature-sensitive lethal region.G3 (Bethesda). 2023 Jun 1;13(6):jkad074. doi: 10.1093/g3journal/jkad074. G3 (Bethesda). 2023. PMID: 36988332 Free PMC article.
-
Identification and functional characterization of major gene pcmfs, controlling cap color formation in Pleurotus cornucopiae.Appl Environ Microbiol. 2025 Mar 19;91(3):e0189424. doi: 10.1128/aem.01894-24. Epub 2025 Feb 27. Appl Environ Microbiol. 2025. PMID: 40013787 Free PMC article.
-
In toto light sheet fluorescence microscopy live imaging datasets of Ceratitis capitata embryonic development.Sci Data. 2022 Jun 15;9(1):340. doi: 10.1038/s41597-022-01443-x. Sci Data. 2022. PMID: 35705572 Free PMC article.
-
A self-limiting sterile insect technique alternative for Ceratitis capitata.BMC Biol. 2025 Apr 12;23(1):97. doi: 10.1186/s12915-025-02201-2. BMC Biol. 2025. PMID: 40221789 Free PMC article.
References
-
- Robinson, A. S. & Hooper, G. Fruit Flies: Their Biology, Natural Enemies, and Control Vol. 1 (Elsevier, 1989).
-
- Dyck, V. A. et al. Sterile Insect Technique – Principles and Practice in Area-Wide Integrated Pest Management (eds Dyck, V. A., Hendrichs, J. & Robinson, A. S.) (Springer, 2005).
-
- Vreysen, M., Robinson, A. S. & Hendrichs, J. Area-Wide Control of Insect Pests: from Research to Field Implementation (Springer, 2007).
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
