

Biomolecular condensation of NUP98 fusion proteins drives leukemogenic gene expression
- PMID: 33479542
- PMCID: PMC7116736
- DOI: 10.1038/s41594-020-00550-w
Biomolecular condensation of NUP98 fusion proteins drives leukemogenic gene expression
Abstract
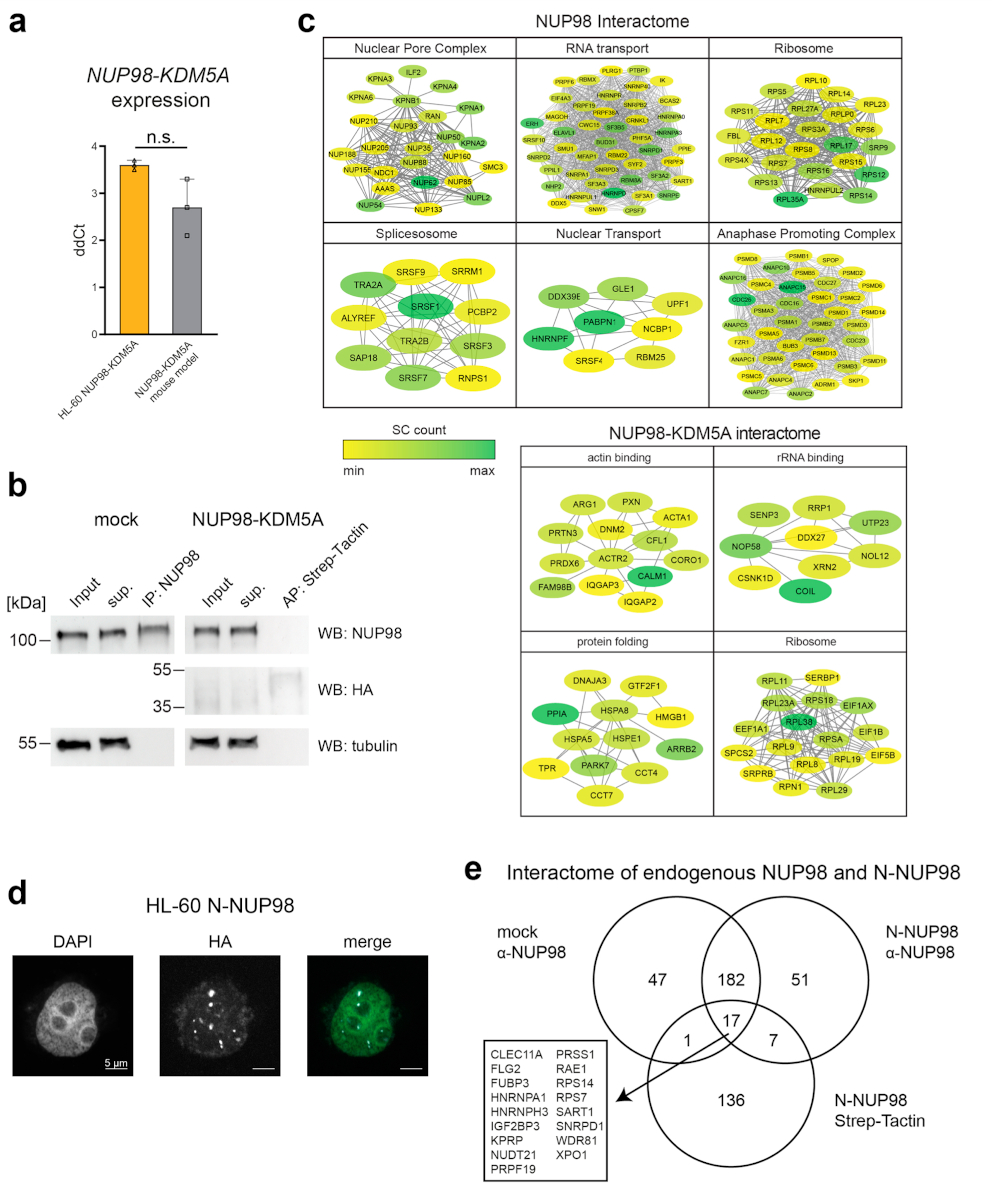
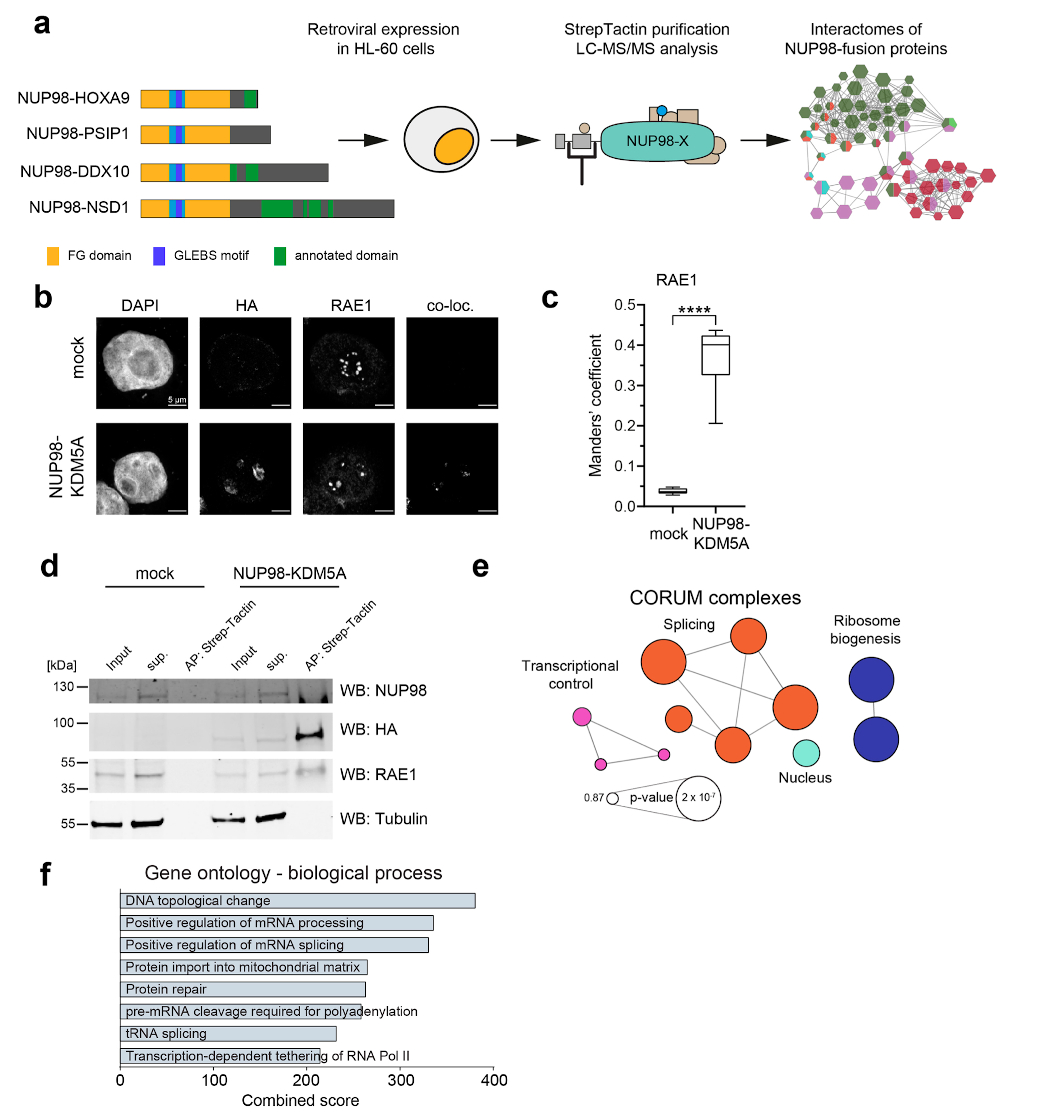
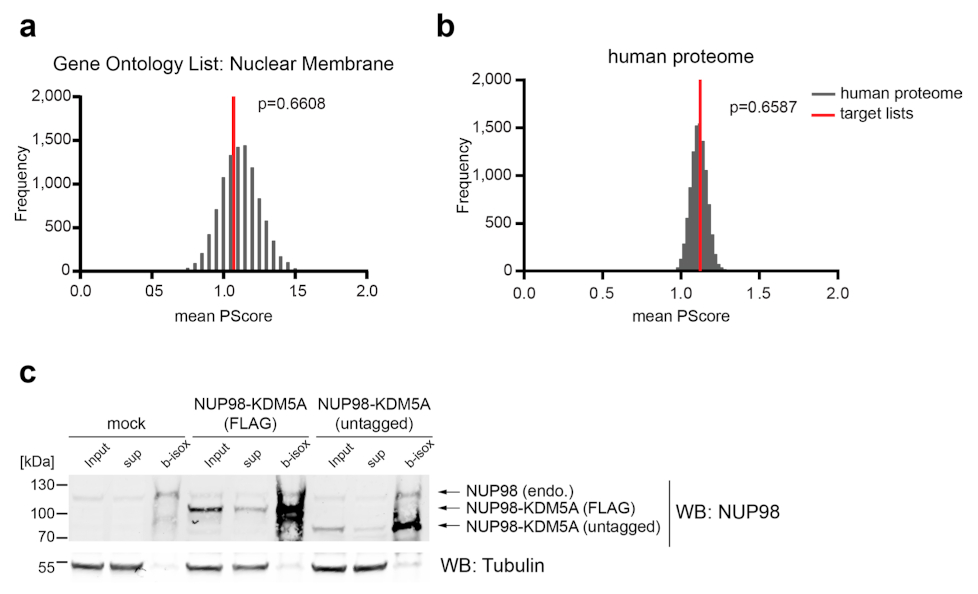
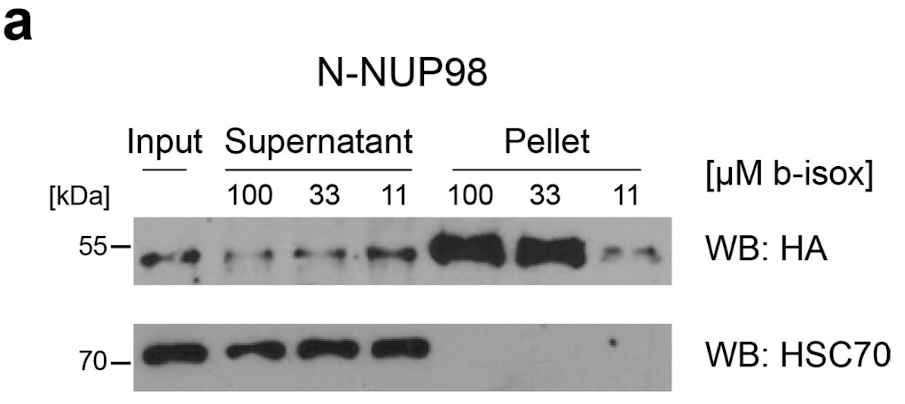
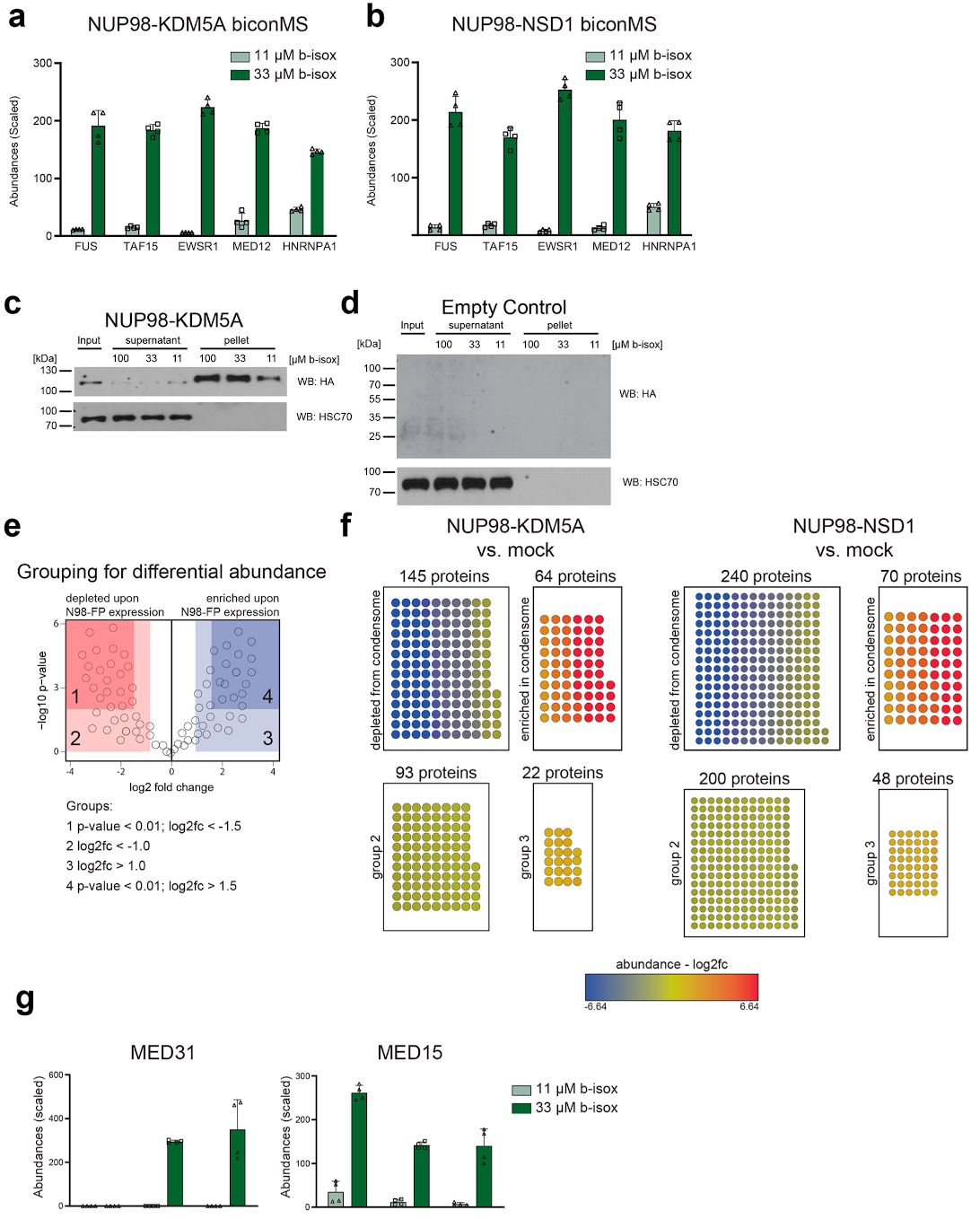
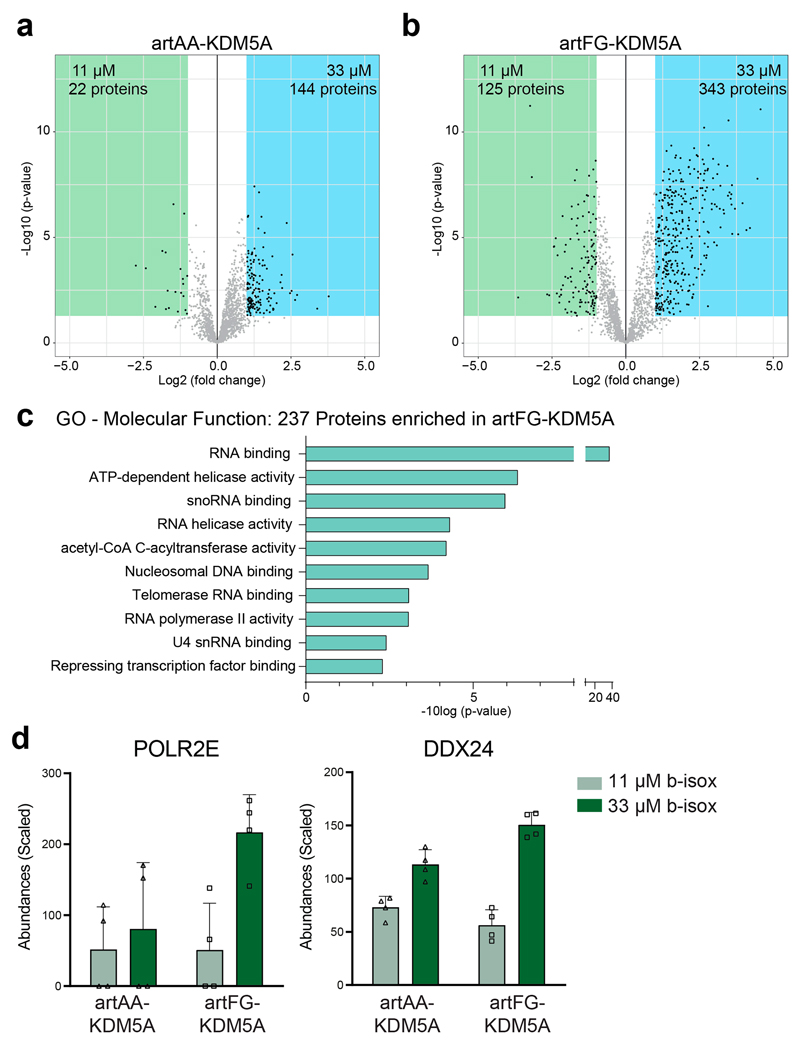
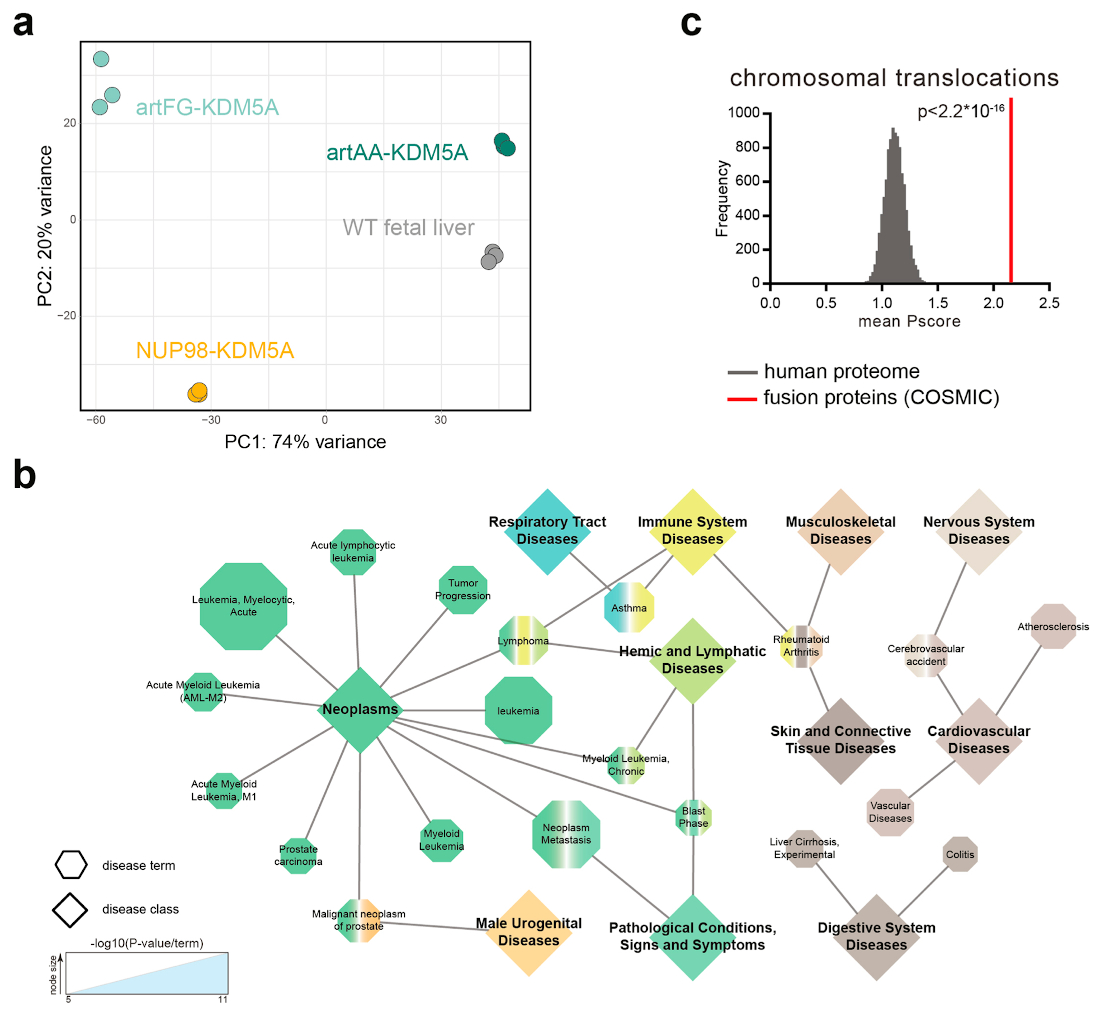
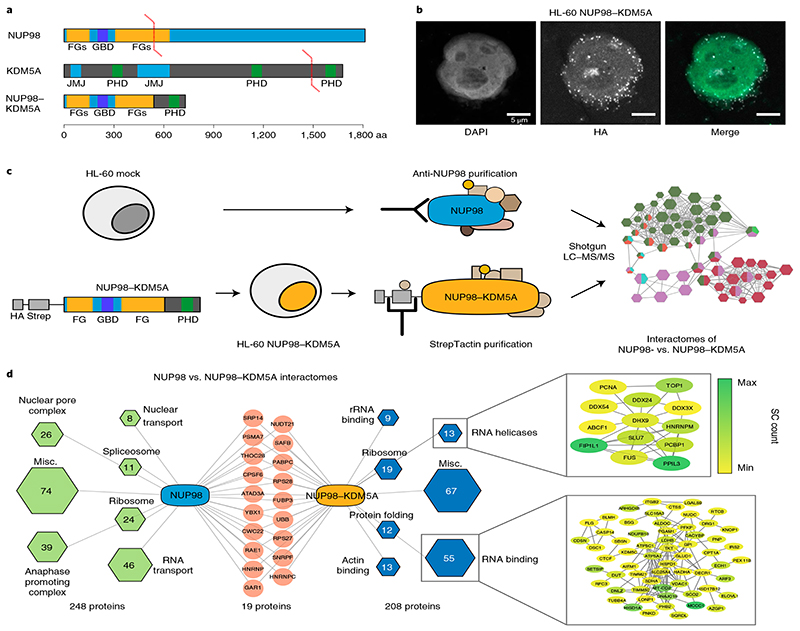
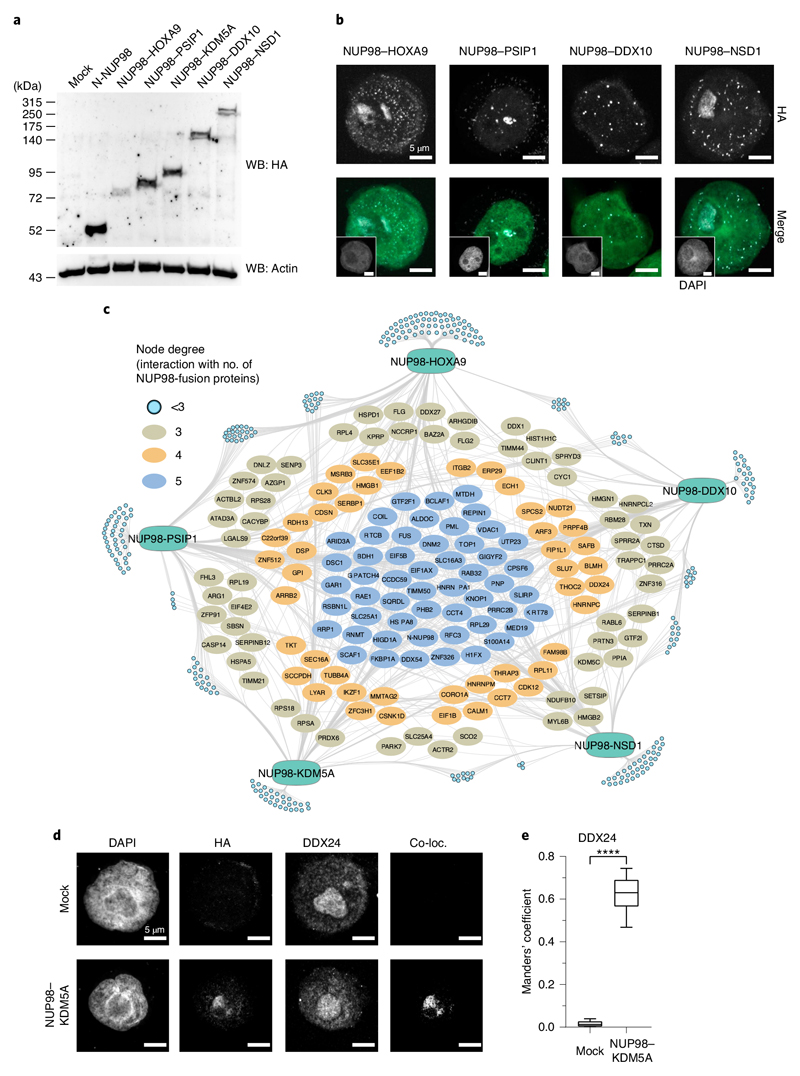
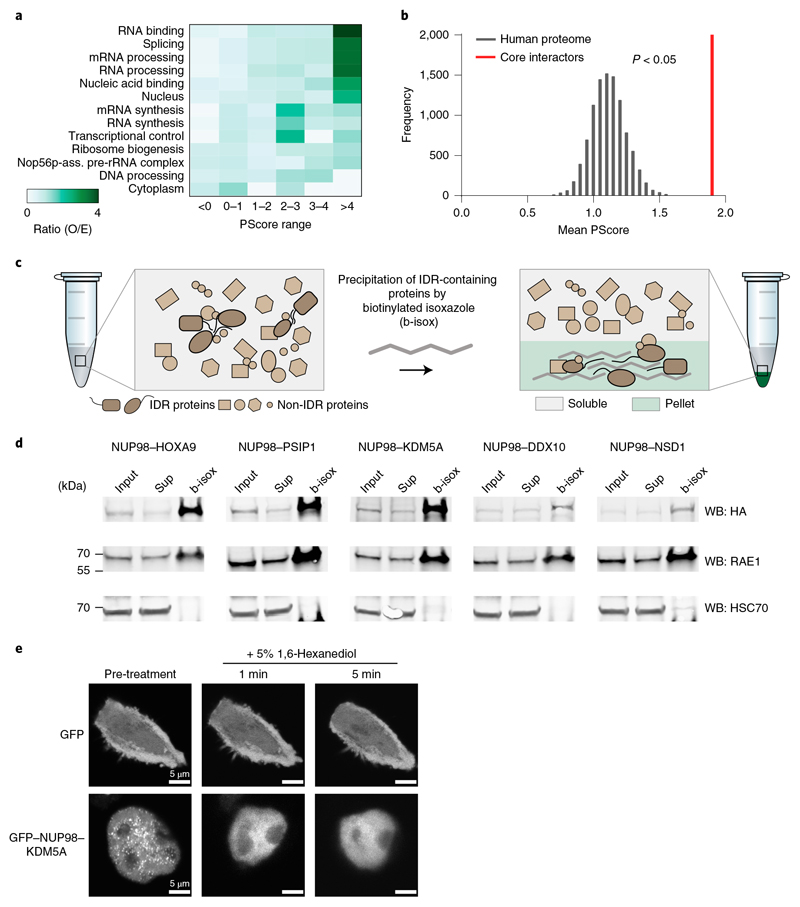
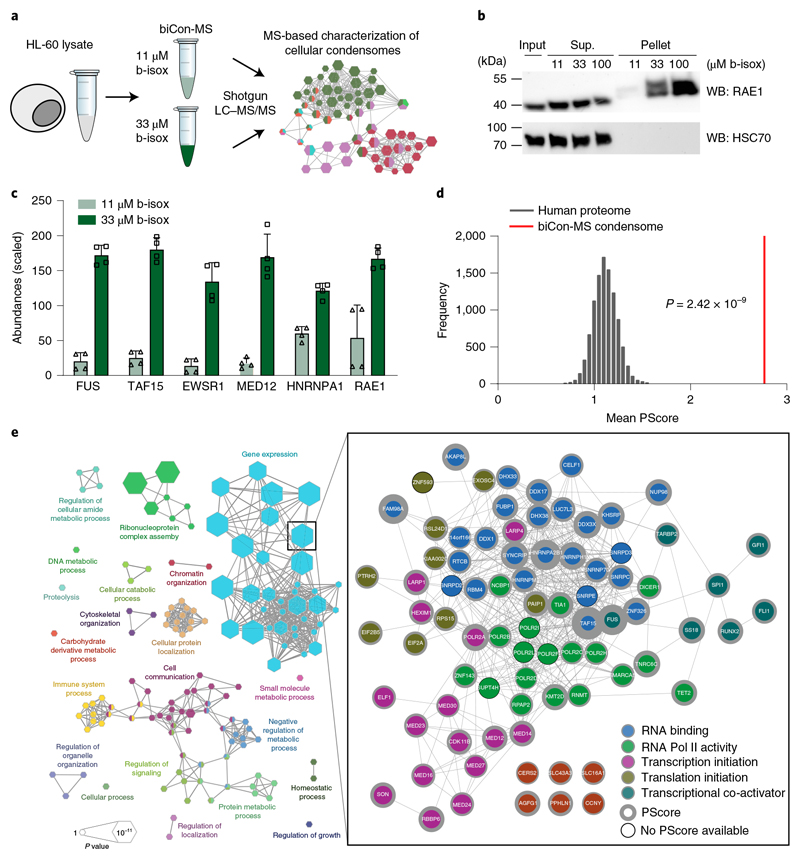
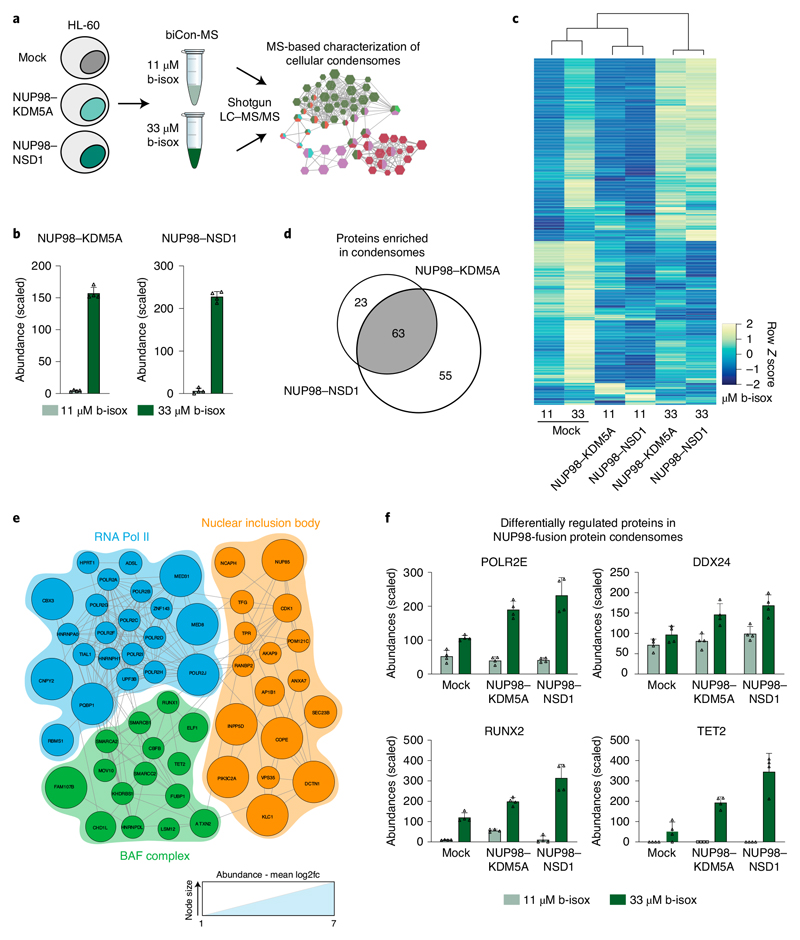
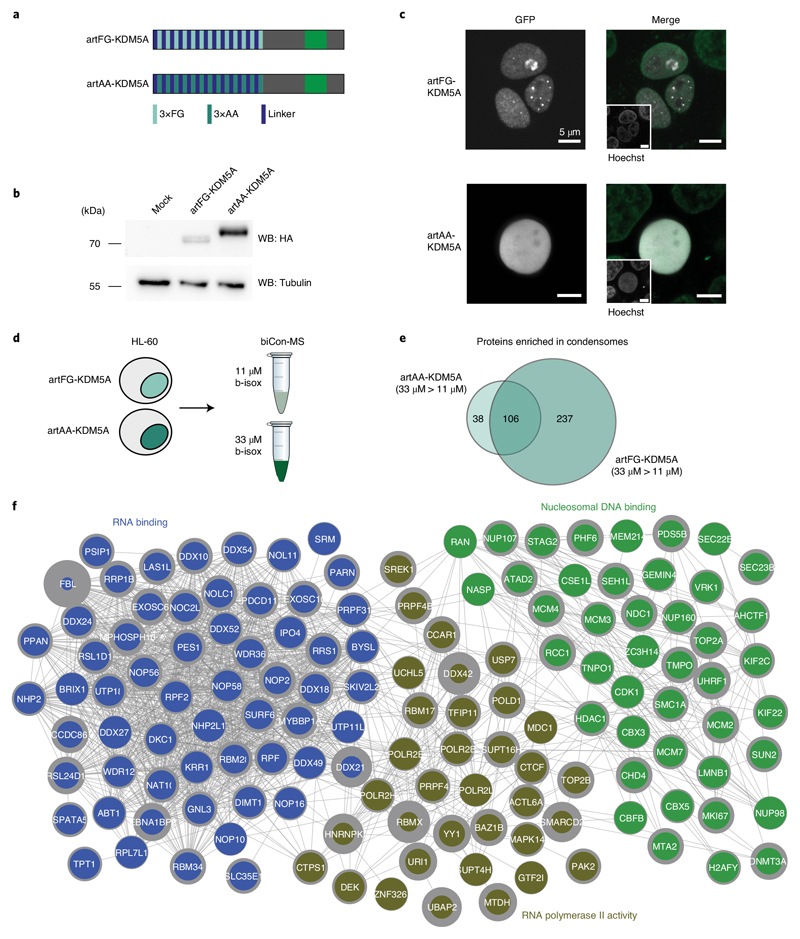
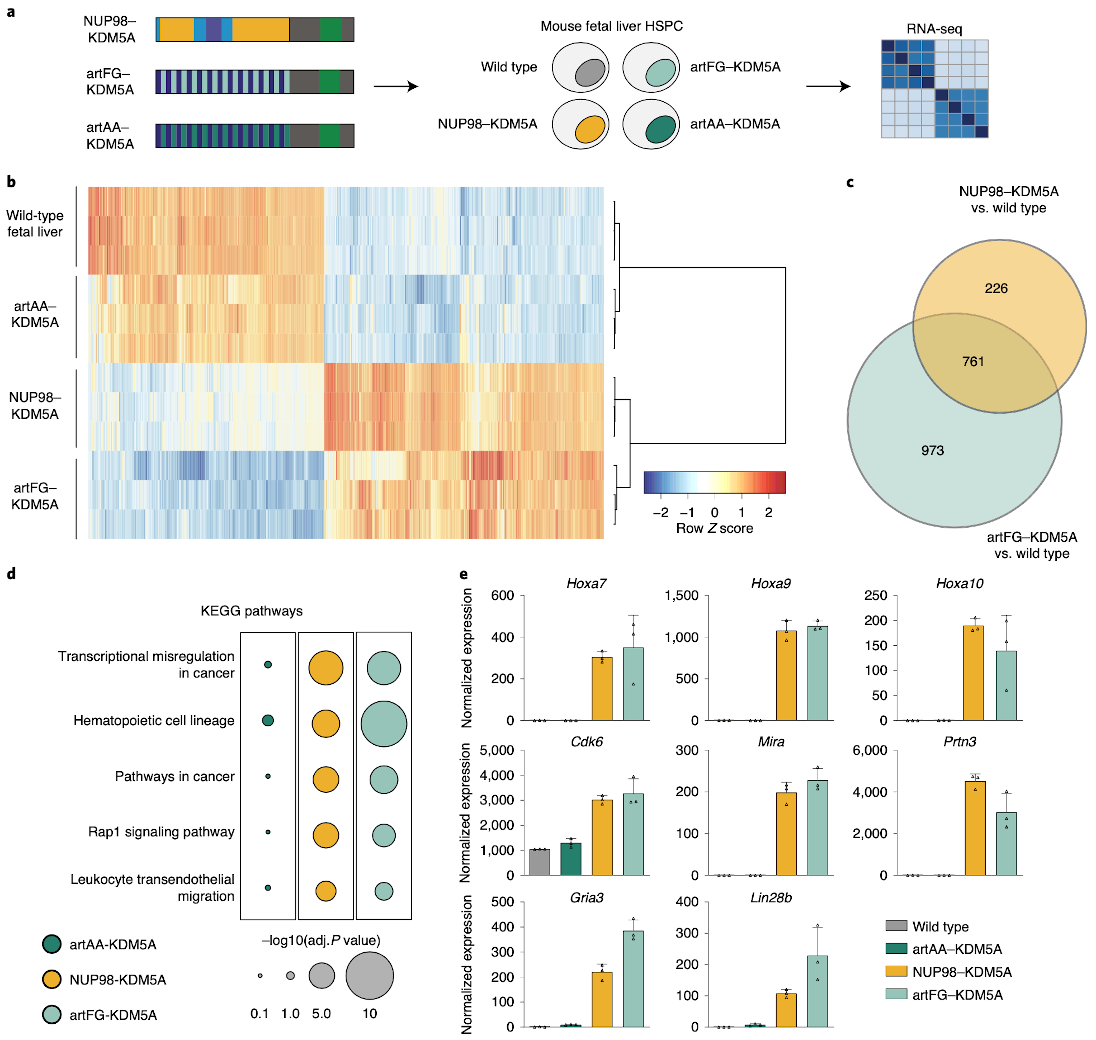
NUP98 fusion proteins cause leukemia via unknown molecular mechanisms. All NUP98 fusion proteins share an intrinsically disordered region (IDR) in the NUP98 N terminus, featuring repeats of phenylalanine-glycine (FG), and C-terminal fusion partners often function in gene control. We investigated whether mechanisms of oncogenic transformation by NUP98 fusion proteins are hardwired in their protein interactomes. Affinity purification coupled to mass spectrometry (MS) and confocal imaging of five NUP98 fusion proteins expressed in human leukemia cells revealed that shared interactors were enriched for proteins involved in biomolecular condensation and that they colocalized with NUP98 fusion proteins in nuclear puncta. We developed biotinylated isoxazole-mediated condensome MS (biCon-MS) to show that NUP98 fusion proteins alter the global composition of biomolecular condensates. An artificial FG-repeat-containing fusion protein phenocopied the nuclear localization patterns of NUP98 fusion proteins and their capability to drive oncogenic gene expression programs. Thus, we propose that IDR-containing fusion proteins combine biomolecular condensation with transcriptional control to induce cancer.
Conflict of interest statement
The authors declare no competing interests.
Figures
Comment in
-
Seeking commonalities in fusion proteins of NUP98.Nat Struct Mol Biol. 2021 Feb;28(2):123-124. doi: 10.1038/s41594-021-00558-w. Nat Struct Mol Biol. 2021. PMID: 33479543 No abstract available.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
