

Seipin traps triacylglycerols to facilitate their nanoscale clustering in the endoplasmic reticulum membrane
- PMID: 33481779
- PMCID: PMC7857593
- DOI: 10.1371/journal.pbio.3000998
Seipin traps triacylglycerols to facilitate their nanoscale clustering in the endoplasmic reticulum membrane
Abstract
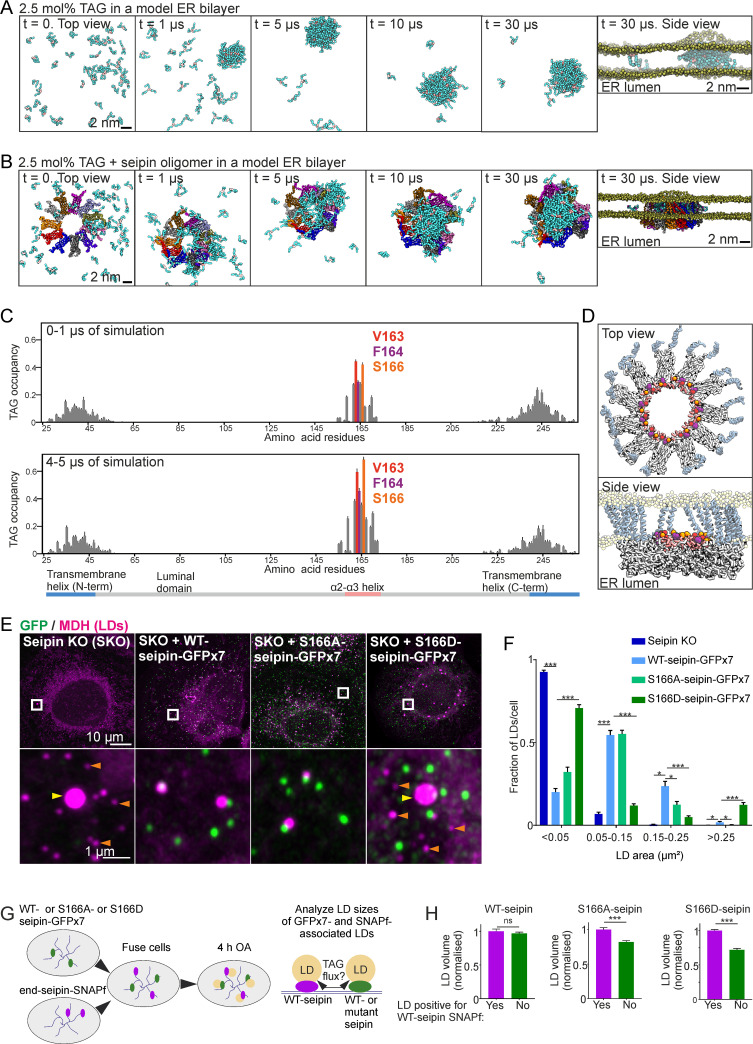
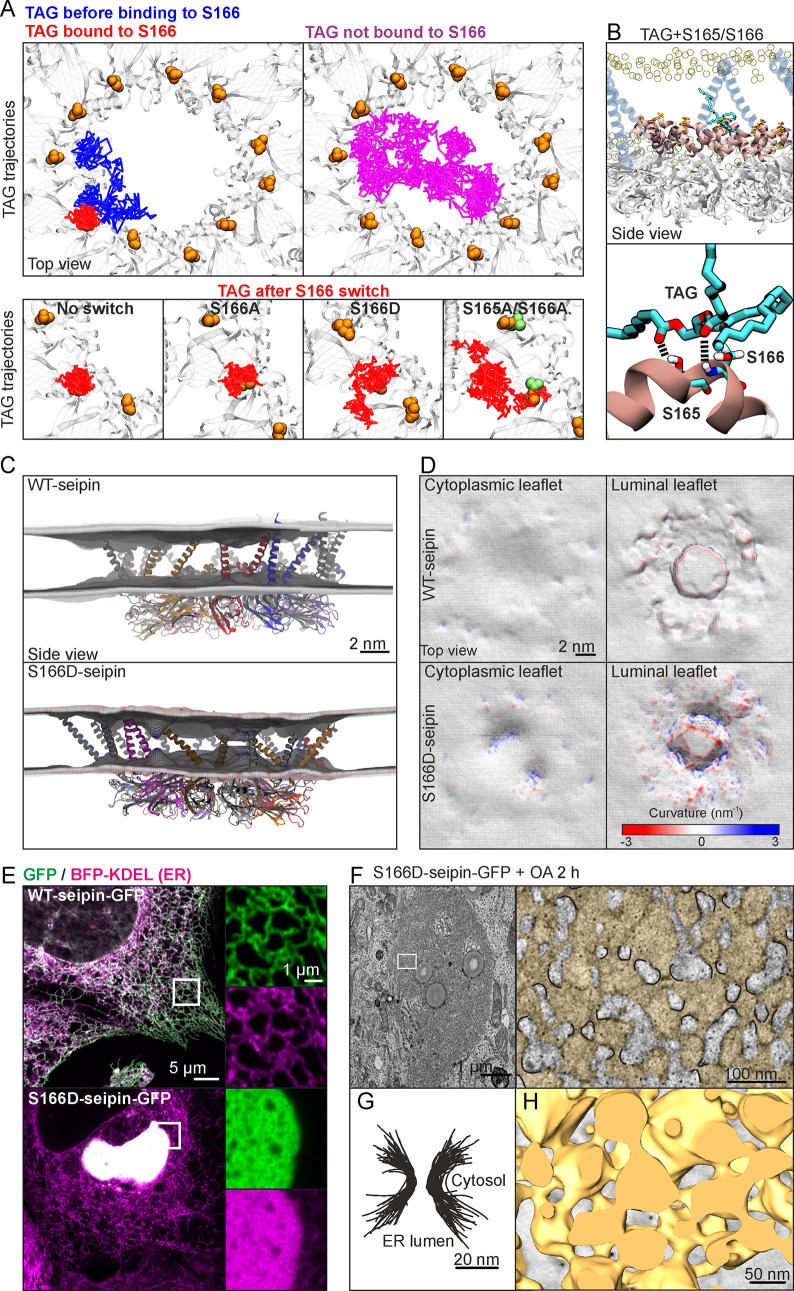
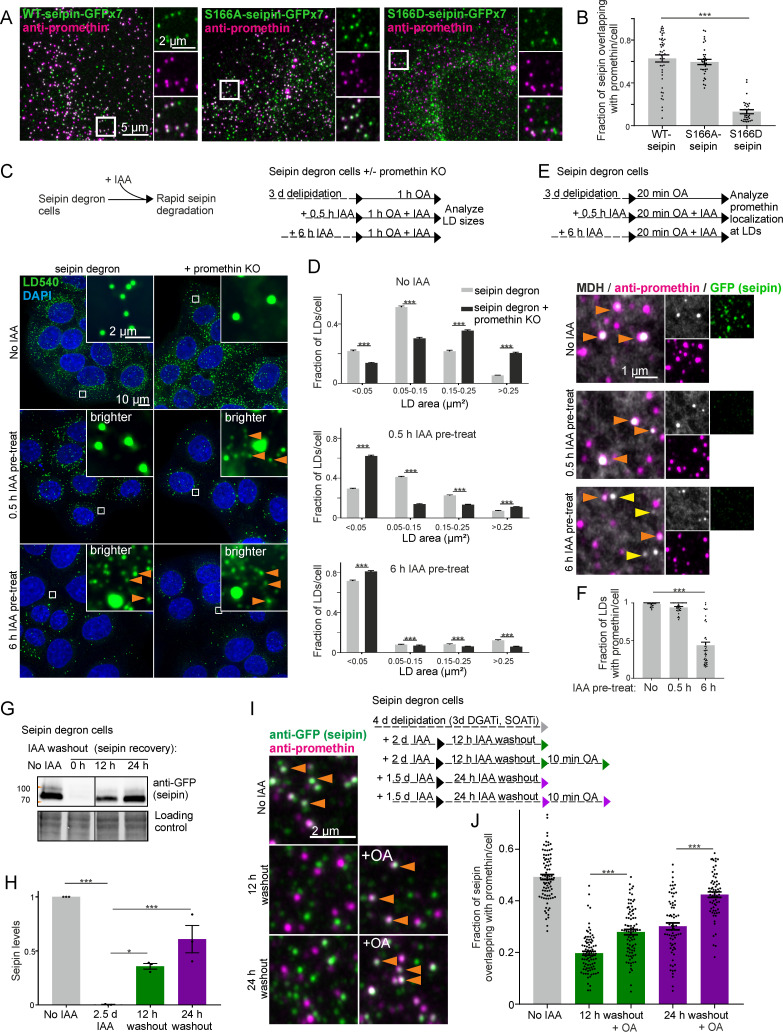
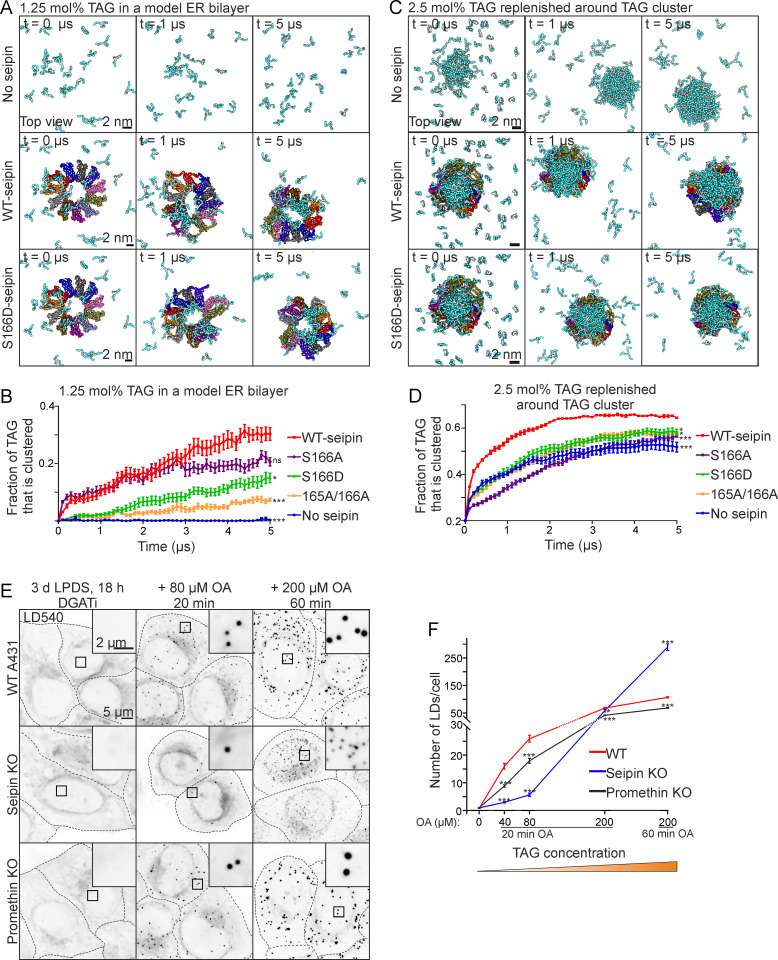
Seipin is a disk-like oligomeric endoplasmic reticulum (ER) protein important for lipid droplet (LD) biogenesis and triacylglycerol (TAG) delivery to growing LDs. Here we show through biomolecular simulations bridged to experiments that seipin can trap TAGs in the ER bilayer via the luminal hydrophobic helices of the protomers delineating the inner opening of the seipin disk. This promotes the nanoscale sequestration of TAGs at a concentration that by itself is insufficient to induce TAG clustering in a lipid membrane. We identify Ser166 in the α3 helix as a favored TAG occupancy site and show that mutating it compromises the ability of seipin complexes to sequester TAG in silico and to promote TAG transfer to LDs in cells. While the S166D-seipin mutant colocalizes poorly with promethin, the association of nascent wild-type seipin complexes with promethin is promoted by TAGs. Together, these results suggest that seipin traps TAGs via its luminal hydrophobic helices, serving as a catalyst for seeding the TAG cluster from dissolved monomers inside the seipin ring, thereby generating a favorable promethin binding interface.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
