

ER/K-link-Leveraging a native protein linker to probe dynamic cellular interactions
- PMID: 33482988
- PMCID: PMC8009693
- DOI: 10.1016/bs.mie.2020.10.002
ER/K-link-Leveraging a native protein linker to probe dynamic cellular interactions
Abstract
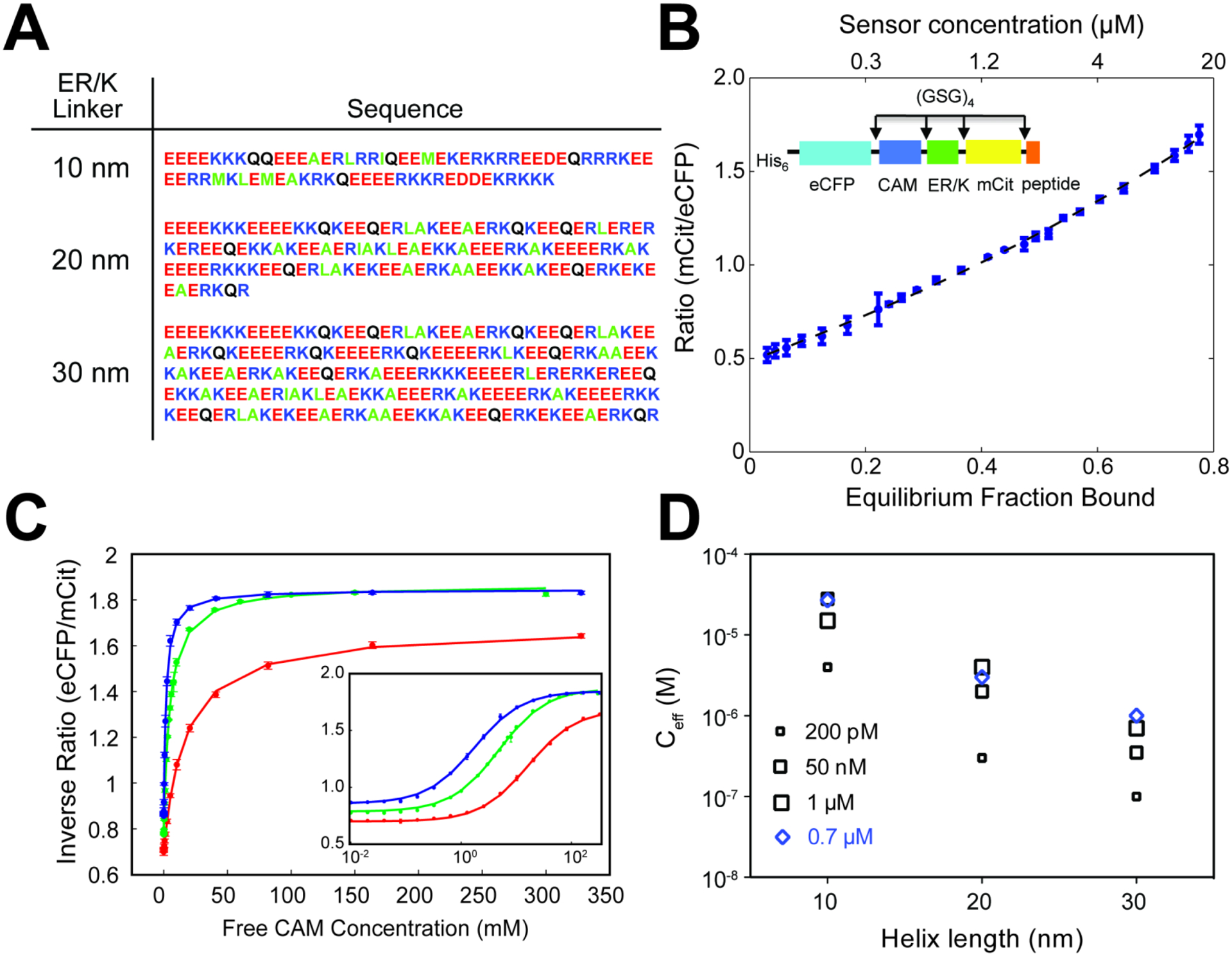
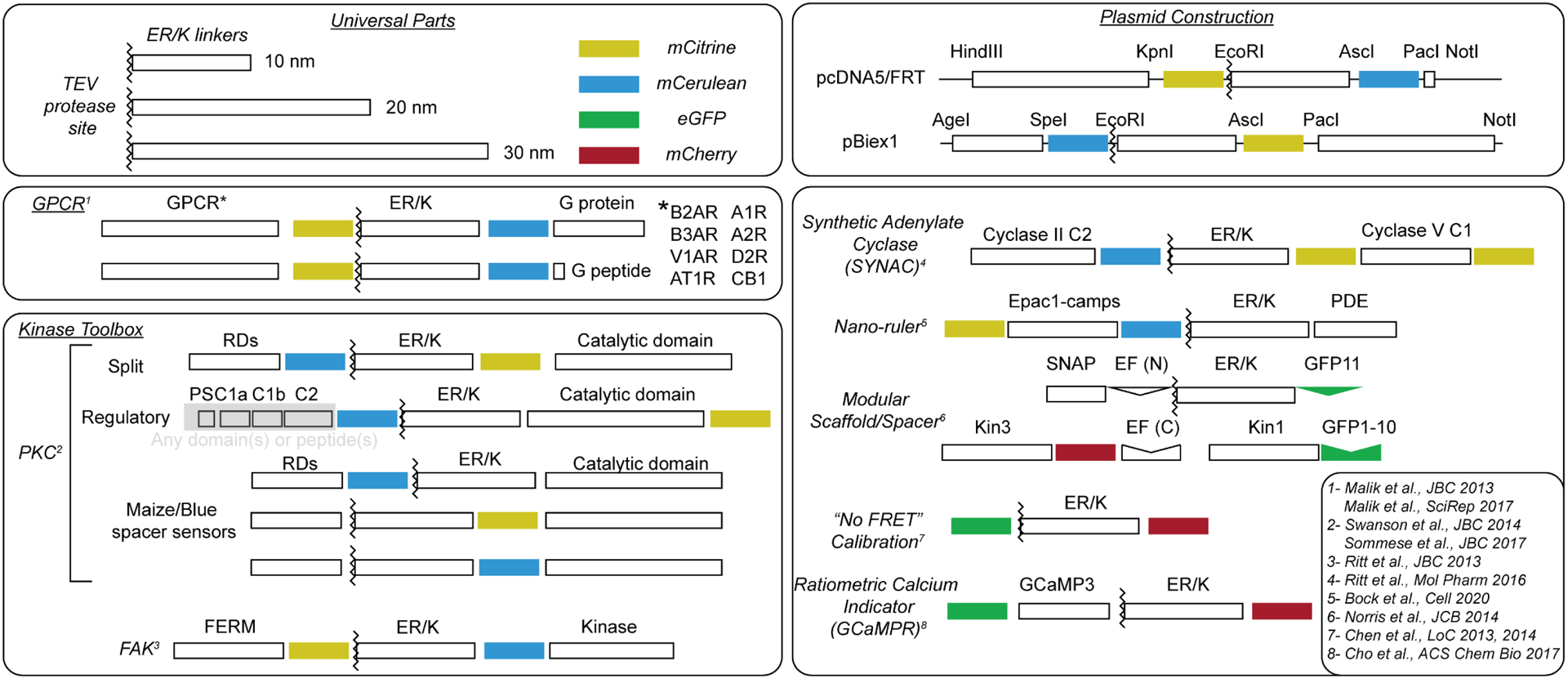
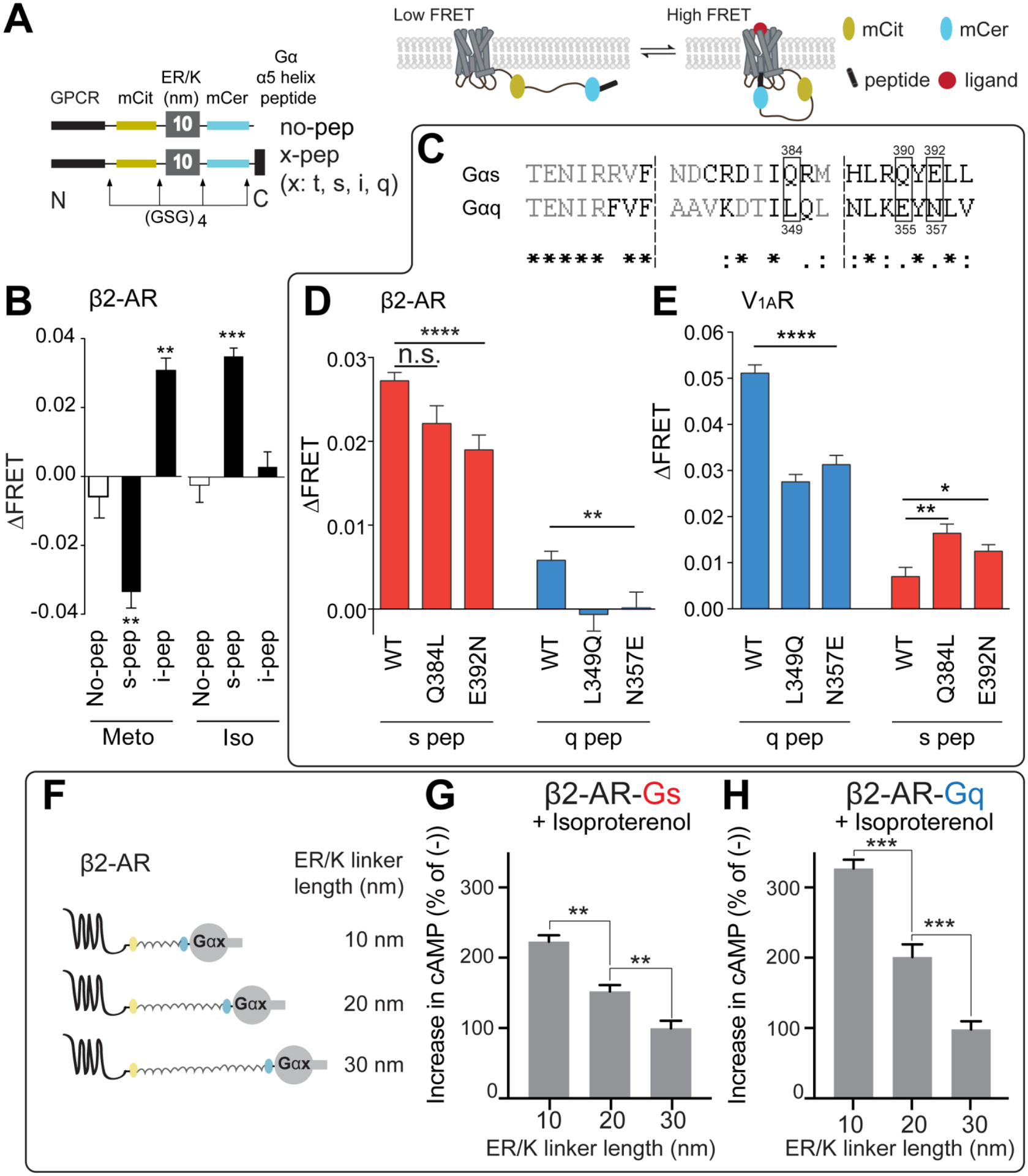
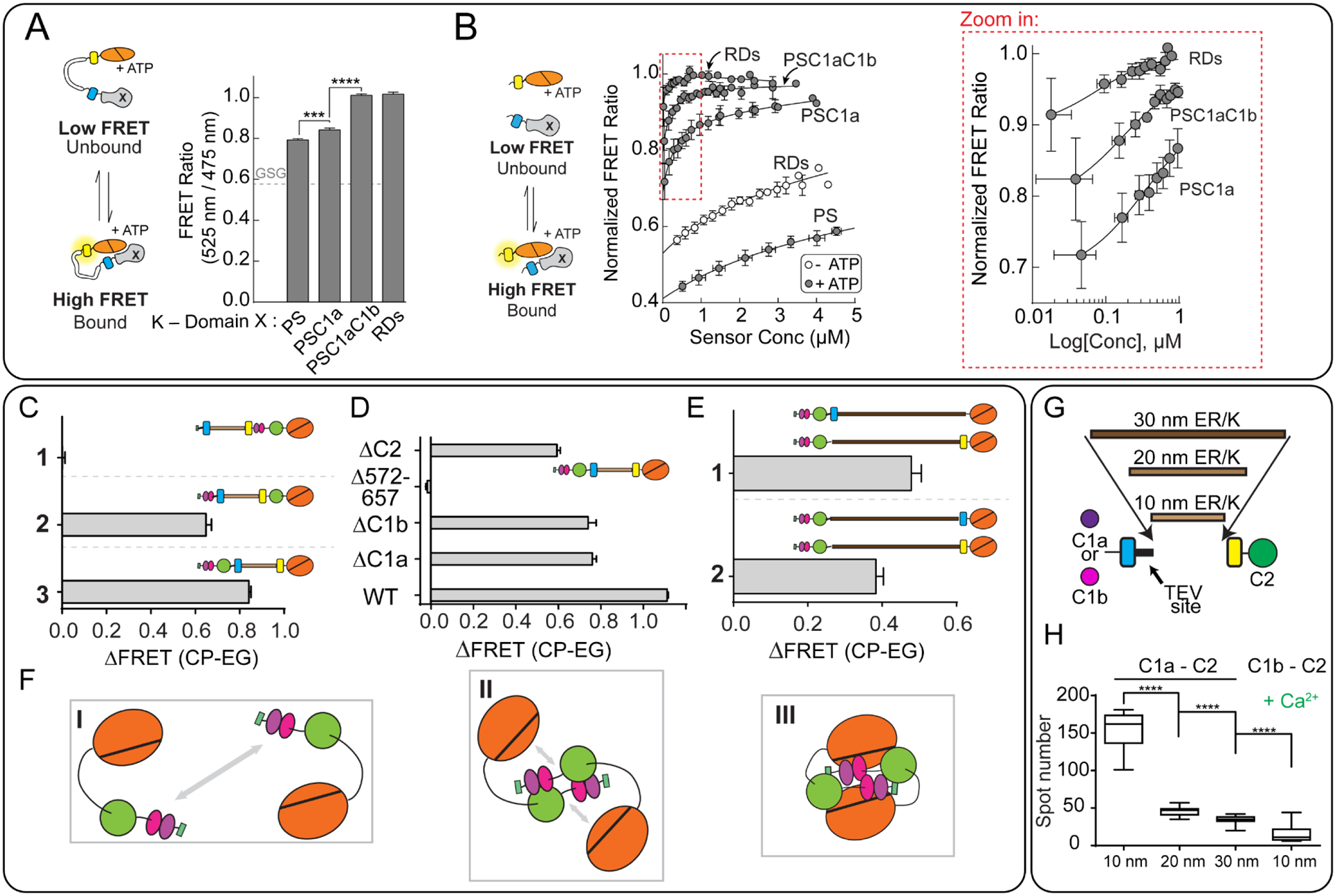
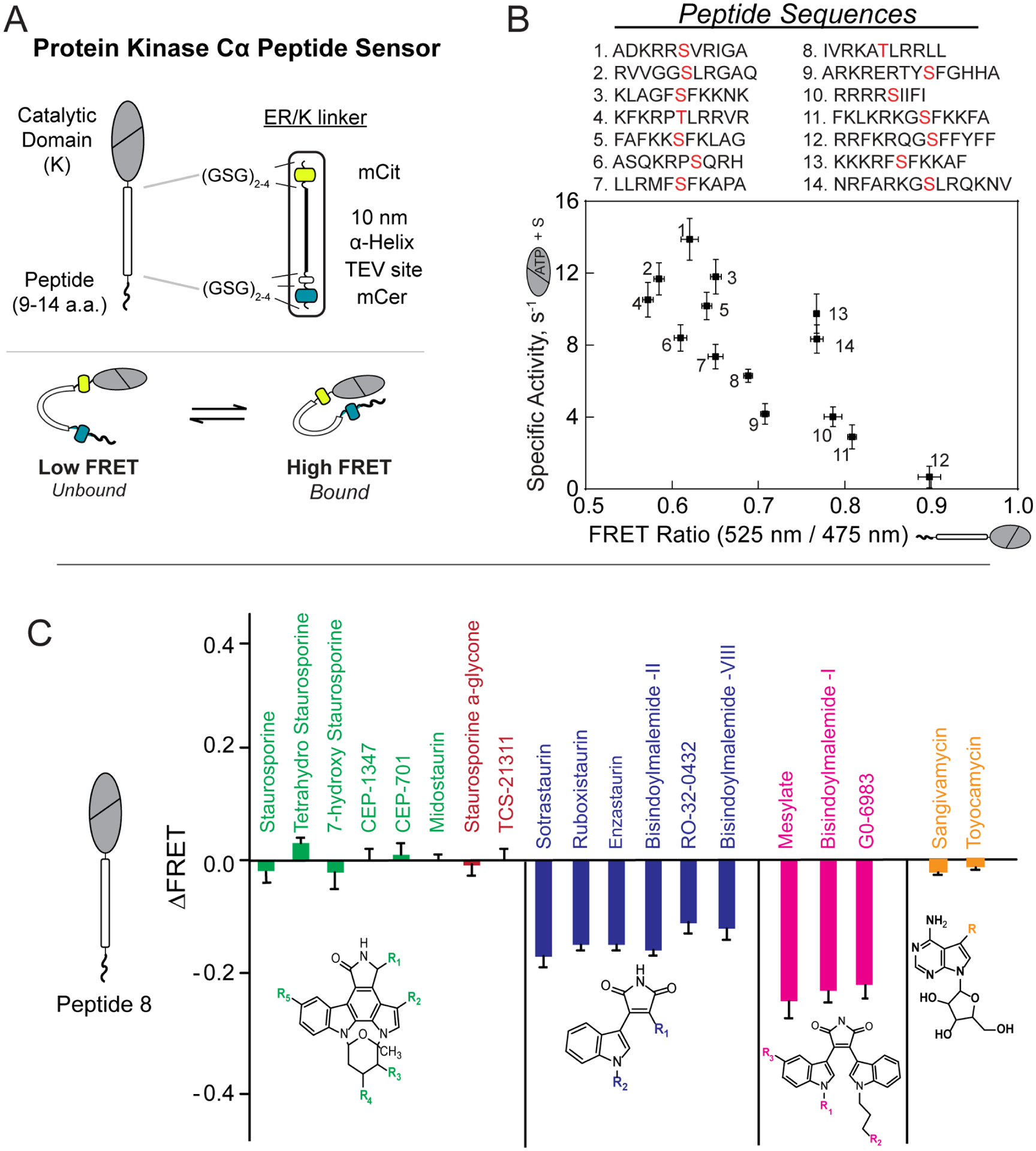
ER/K α-helices are a subset of single alpha helical domains, which exhibit unusual stability as isolated protein secondary structures. They adopt an elongated structural conformation, while regulating the frequency of interactions between proteins or polypeptides fused to their ends. Here we review recent advances on the structure, stability and function of ER/K α-helices as linkers (ER/K linkers) in native proteins. We describe methodological considerations in the molecular cloning, protein expression and measurement of interaction strengths, using sensors incorporating ER/K linkers. We highlight biological insights obtained over the last decade by leveraging distinct biophysical features of ER/K-linked sensors. We conclude with the outlook for the use of ER/K linkers in the selective modulation of dynamic cellular interactions.
Keywords: ER/K linker; FRET biosensors; GPCR; Kinases; Protein-protein interactions; Signaling; Single alpha helix.
© 2021 Elsevier Inc. All rights reserved.
Figures
Similar articles
-
Harnessing the unique structural properties of isolated α-helices.J Biol Chem. 2014 Sep 12;289(37):25460-7. doi: 10.1074/jbc.R114.583906. Epub 2014 Jul 24. J Biol Chem. 2014. PMID: 25059657 Free PMC article.
-
Systematic control of protein interaction using a modular ER/K α-helix linker.Proc Natl Acad Sci U S A. 2011 Dec 20;108(51):20467-72. doi: 10.1073/pnas.1116066108. Epub 2011 Nov 28. Proc Natl Acad Sci U S A. 2011. PMID: 22123984 Free PMC article.
-
Dynamic charge interactions create surprising rigidity in the ER/K alpha-helical protein motif.Proc Natl Acad Sci U S A. 2008 Sep 9;105(36):13356-61. doi: 10.1073/pnas.0806256105. Epub 2008 Sep 3. Proc Natl Acad Sci U S A. 2008. PMID: 18768817 Free PMC article.
-
Design of helical linkers for fusion proteins and protein-based nanostructures.Methods Enzymol. 2021;647:209-230. doi: 10.1016/bs.mie.2020.10.003. Epub 2020 Nov 18. Methods Enzymol. 2021. PMID: 33482989 Review.
-
The linker of calmodulin--to helix or not to helix.Cell Calcium. 1992 Jun-Jul;13(6-7):363-76. doi: 10.1016/0143-4160(92)90050-3. Cell Calcium. 1992. PMID: 1505002 Review.
Cited by
-
Advancements in G protein-coupled receptor biosensors to study GPCR-G protein coupling.Br J Pharmacol. 2023 Jun;180(11):1433-1443. doi: 10.1111/bph.15962. Epub 2022 Oct 17. Br J Pharmacol. 2023. PMID: 36166832 Free PMC article. Review.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
