

Conjugative plasmid-encoded toxin-antitoxin system PrpT/PrpA directly controls plasmid copy number
- PMID: 33483419
- PMCID: PMC7848731
- DOI: 10.1073/pnas.2011577118
Conjugative plasmid-encoded toxin-antitoxin system PrpT/PrpA directly controls plasmid copy number
Abstract
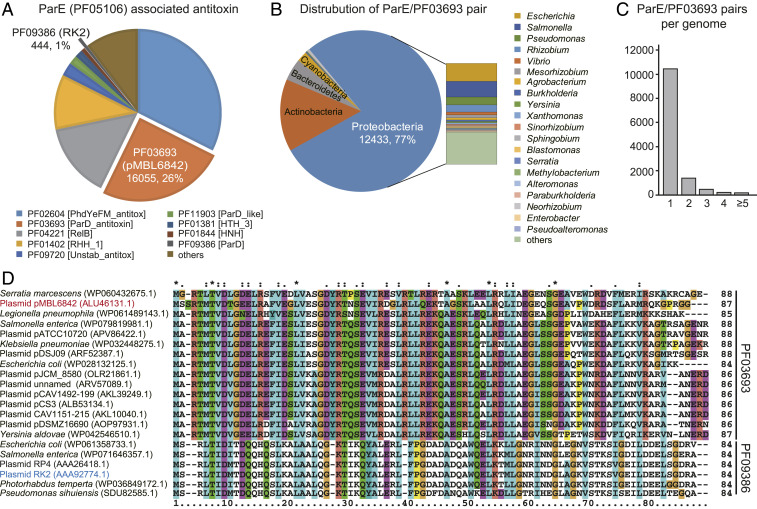
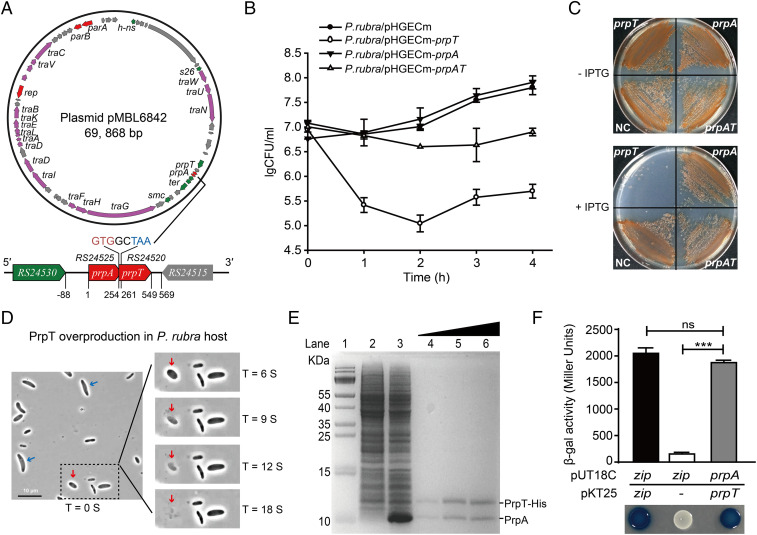
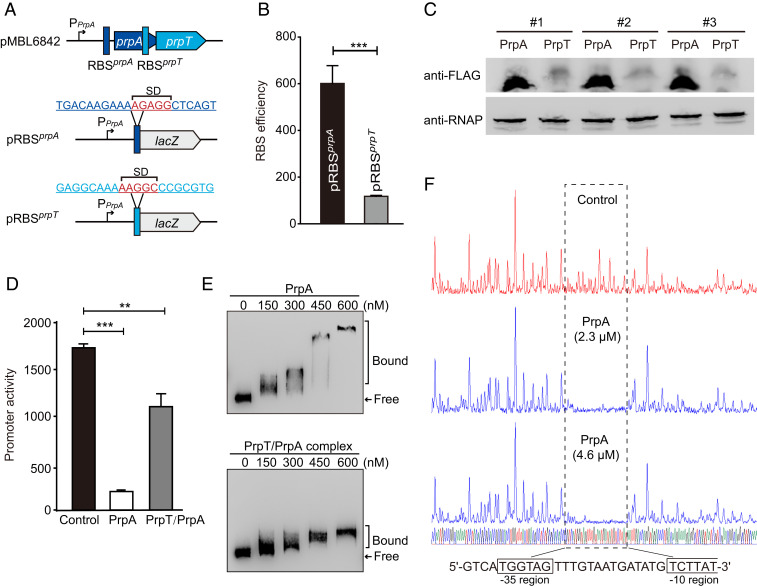
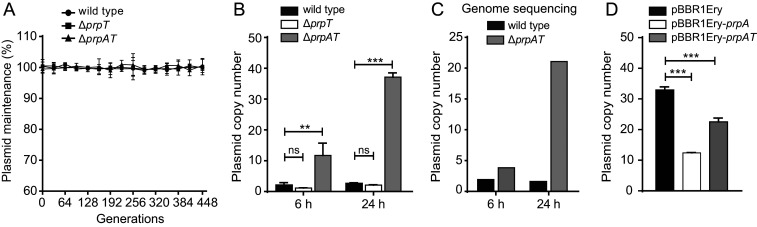
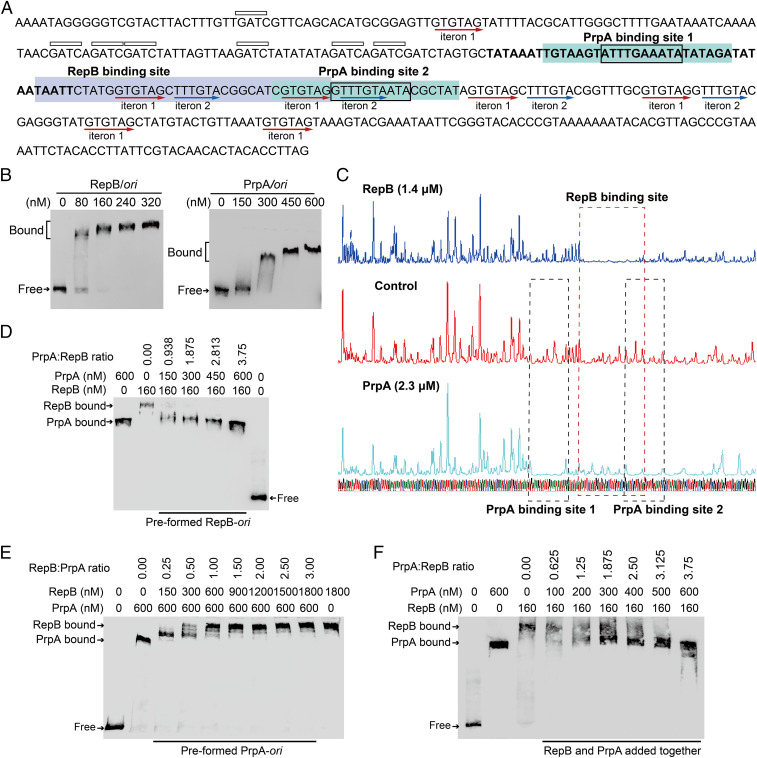
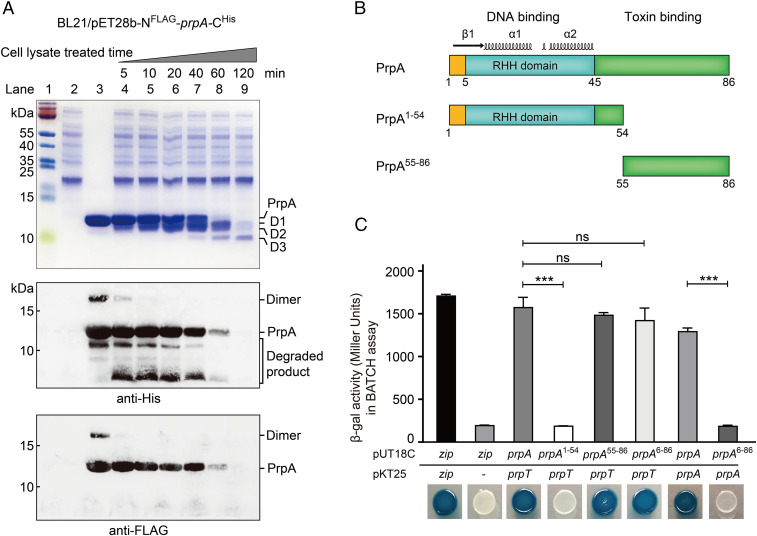
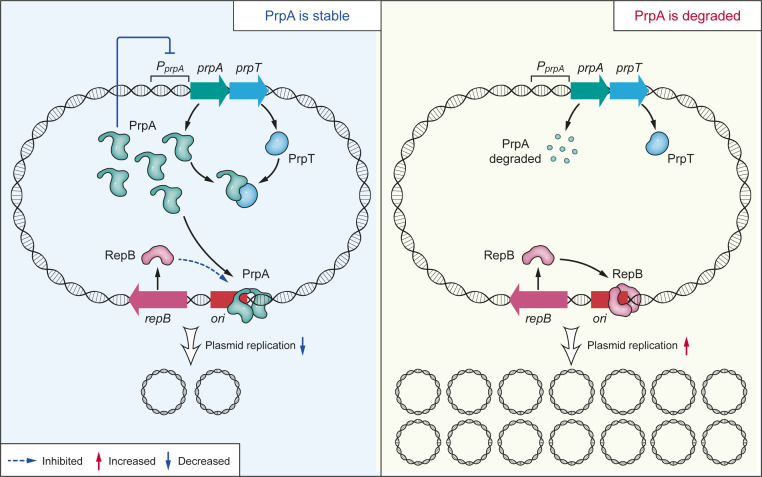
Toxin-antitoxin (TA) loci were initially identified on conjugative plasmids, and one function of plasmid-encoded TA systems is to stabilize plasmids or increase plasmid competition via postsegregational killing. Here, we discovered that the type II TA system, Pseudoalteromonas rubra plasmid toxin-antitoxin PrpT/PrpA, on a low-copy-number conjugative plasmid, directly controls plasmid replication. Toxin PrpT resembles ParE of plasmid RK2 while antitoxin PrpA (PF03693) shares no similarity with previously characterized antitoxins. Surprisingly, deleting this prpA-prpT operon from the plasmid does not result in plasmid segregational loss, but greatly increases plasmid copy number. Mechanistically, the antitoxin PrpA functions as a negative regulator of plasmid replication, by binding to the iterons in the plasmid origin that inhibits the binding of the replication initiator to the iterons. We also demonstrated that PrpA is produced at a higher level than PrpT to prevent the plasmid from overreplicating, while partial or complete degradation of labile PrpA derepresses plasmid replication. Importantly, the PrpT/PrpA TA system is conserved and is widespread on many conjugative plasmids. Altogether, we discovered a function of a plasmid-encoded TA system that provides new insights into the physiological significance of TA systems.
Keywords: ParE; origin of replication; plasmid copy number; plasmid replication; toxin–antitoxin.
Copyright © 2021 the Author(s). Published by PNAS.
Conflict of interest statement
The authors declare no competing interest.
Figures
Similar articles
-
Structural insights into the PrpTA toxin-antitoxin system in Pseudoalteromonas rubra.Front Microbiol. 2022 Nov 24;13:1053255. doi: 10.3389/fmicb.2022.1053255. eCollection 2022. Front Microbiol. 2022. PMID: 36504814 Free PMC article.
-
A noncanonical intrinsic terminator in the HicAB toxin-antitoxin operon promotes the transmission of conjugative antibiotic resistance plasmids.Nucleic Acids Res. 2025 Feb 27;53(5):gkaf125. doi: 10.1093/nar/gkaf125. Nucleic Acids Res. 2025. PMID: 40036506 Free PMC article.
-
Proteolysis in plasmid DNA stable maintenance in bacterial cells.Plasmid. 2016 Jul;86:7-13. doi: 10.1016/j.plasmid.2016.05.002. Epub 2016 May 29. Plasmid. 2016. PMID: 27252071 Review.
-
The higBA-Type Toxin-Antitoxin System in IncC Plasmids Is a Mobilizable Ciprofloxacin-Inducible System.mSphere. 2021 Jun 30;6(3):e0042421. doi: 10.1128/mSphere.00424-21. Epub 2021 Jun 2. mSphere. 2021. PMID: 34077263 Free PMC article.
-
Proteic toxin-antitoxin, bacterial plasmid addiction systems and their evolution with special reference to the pas system of pTF-FC2.FEMS Microbiol Lett. 1999 Jul 15;176(2):269-77. doi: 10.1111/j.1574-6968.1999.tb13672.x. FEMS Microbiol Lett. 1999. PMID: 10427709 Review.
Cited by
-
Type II Toxin-Antitoxin Systems in Pseudomonas aeruginosa.Toxins (Basel). 2023 Feb 17;15(2):164. doi: 10.3390/toxins15020164. Toxins (Basel). 2023. PMID: 36828478 Free PMC article. Review.
-
TADB 3.0: an updated database of bacterial toxin-antitoxin loci and associated mobile genetic elements.Nucleic Acids Res. 2024 Jan 5;52(D1):D784-D790. doi: 10.1093/nar/gkad962. Nucleic Acids Res. 2024. PMID: 37897352 Free PMC article.
-
Antitoxin CrlA of CrlTA Toxin-Antitoxin System in a Clinical Isolate Pseudomonas aeruginosa Inhibits Lytic Phage Infection.Front Microbiol. 2022 May 10;13:892021. doi: 10.3389/fmicb.2022.892021. eCollection 2022. Front Microbiol. 2022. PMID: 35620101 Free PMC article.
-
Interactions of the Streptococcus pneumoniae Toxin-Antitoxin RelBE Proteins with Their Target DNA.Microorganisms. 2021 Apr 15;9(4):851. doi: 10.3390/microorganisms9040851. Microorganisms. 2021. PMID: 33921033 Free PMC article.
-
The HicAB System: Characteristics and Biological Roles of an Underappreciated Toxin-Antitoxin System.Int J Mol Sci. 2024 Nov 13;25(22):12165. doi: 10.3390/ijms252212165. Int J Mol Sci. 2024. PMID: 39596231 Free PMC article. Review.
References
-
- Heuer H., Smalla K., Plasmids foster diversification and adaptation of bacterial populations in soil. FEMS Microbiol. Rev. 36, 1083–1104 (2012). - PubMed
-
- Garcillán-Barcia M. P., Francia M. V., de la Cruz F., The diversity of conjugative relaxases and its application in plasmid classification. FEMS Microbiol. Rev. 33, 657–687 (2009). - PubMed
-
- Hayes F., The function and organization of plasmids. Methods Mol. Biol. 235, 1–17 (2003). - PubMed
-
- Nordström K., Austin S. J., Mechanisms that contribute to the stable segregation of plasmids. Annu. Rev. Genet. 23, 37–69 (1989). - PubMed
Publication types
MeSH terms
Substances
Supplementary concepts
LinkOut - more resources
Full Text Sources
Other Literature Sources
