

Chromosomal scale assembly of parasitic wasp genome reveals symbiotic virus colonization
- PMID: 33483589
- PMCID: PMC7822920
- DOI: 10.1038/s42003-020-01623-8
Chromosomal scale assembly of parasitic wasp genome reveals symbiotic virus colonization
Erratum in
-
Author Correction: Chromosomal scale assembly of parasitic wasp genome reveals symbiotic virus colonization.Commun Biol. 2021 Jul 30;4(1):940. doi: 10.1038/s42003-021-02480-9. Commun Biol. 2021. PMID: 34331006 Free PMC article. No abstract available.
Abstract
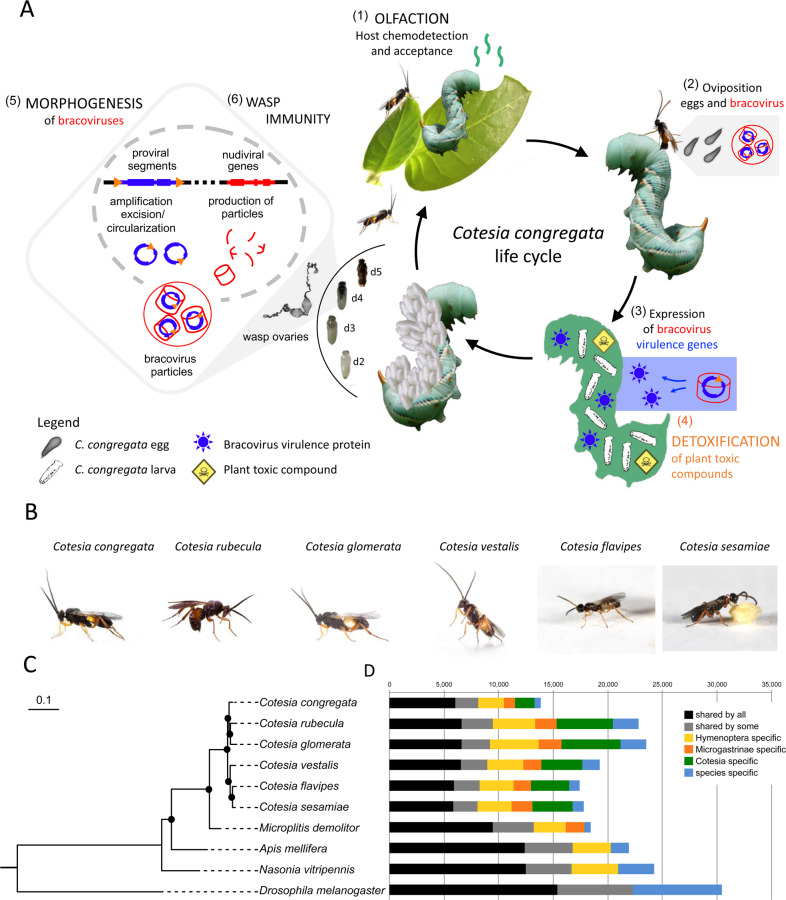
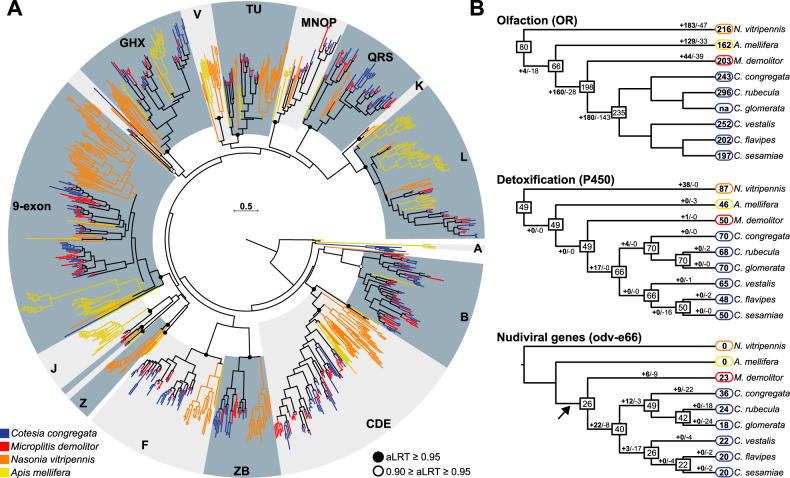
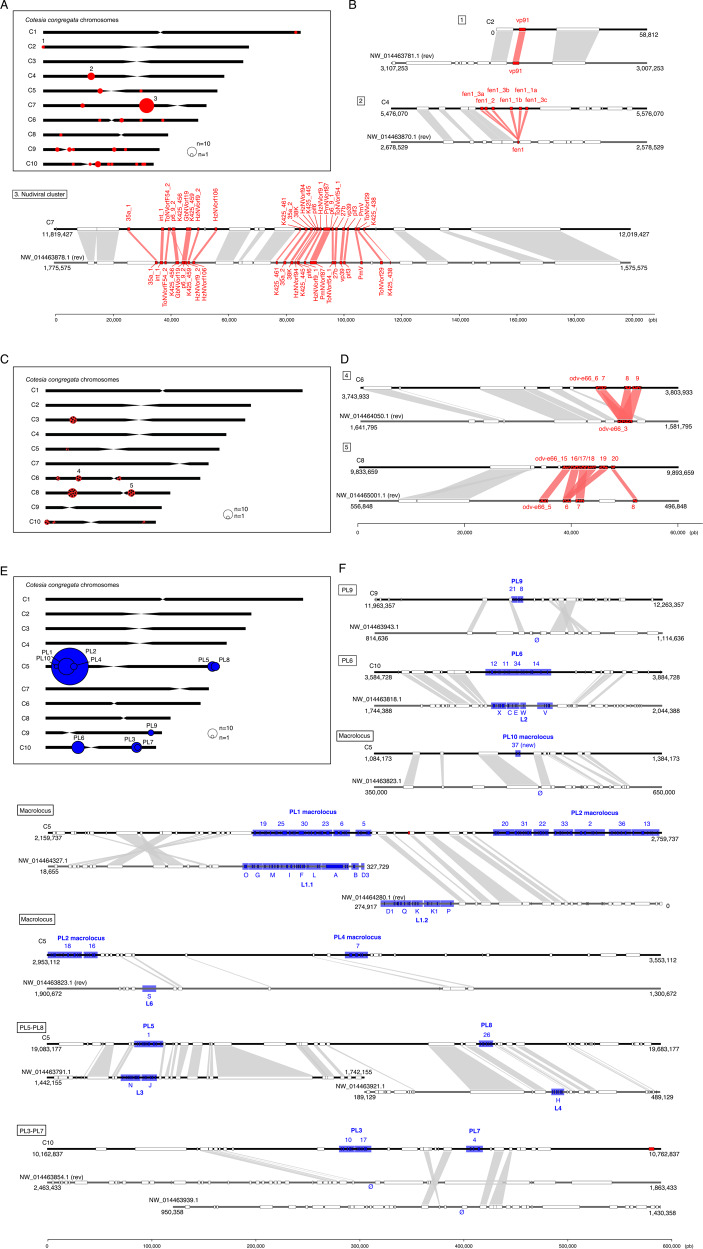
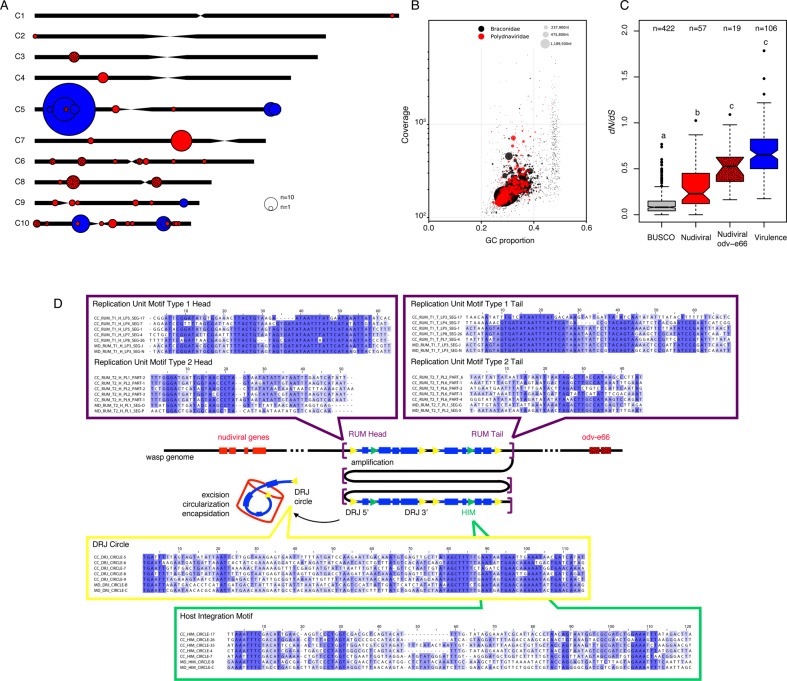
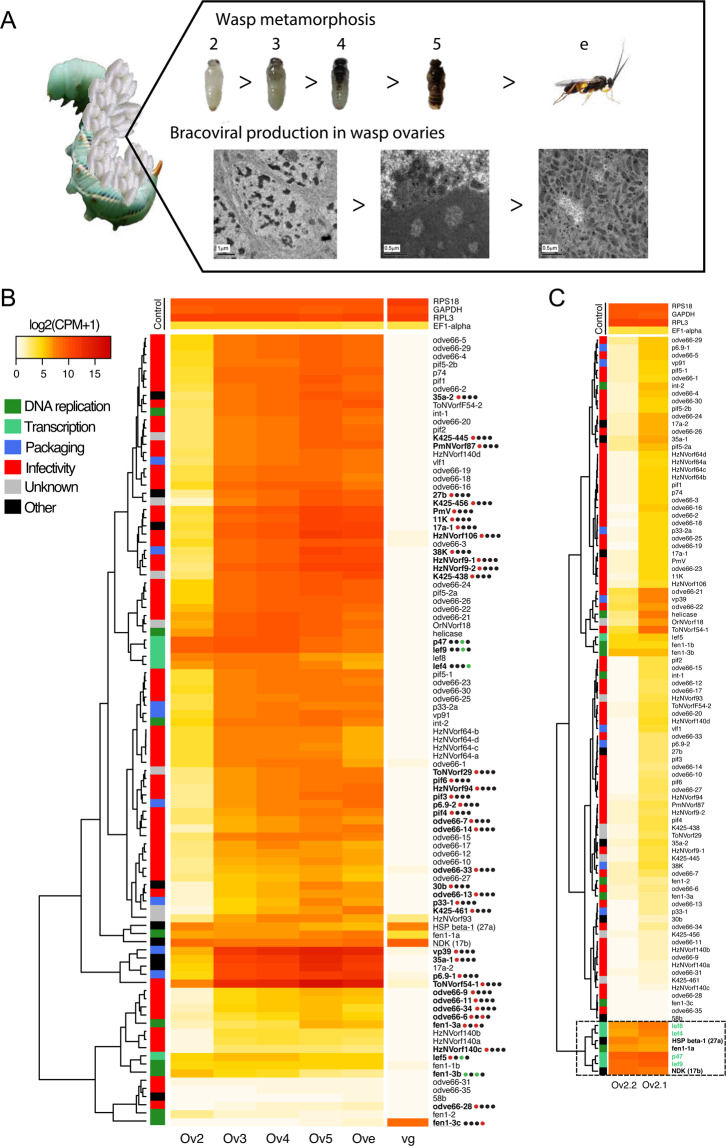
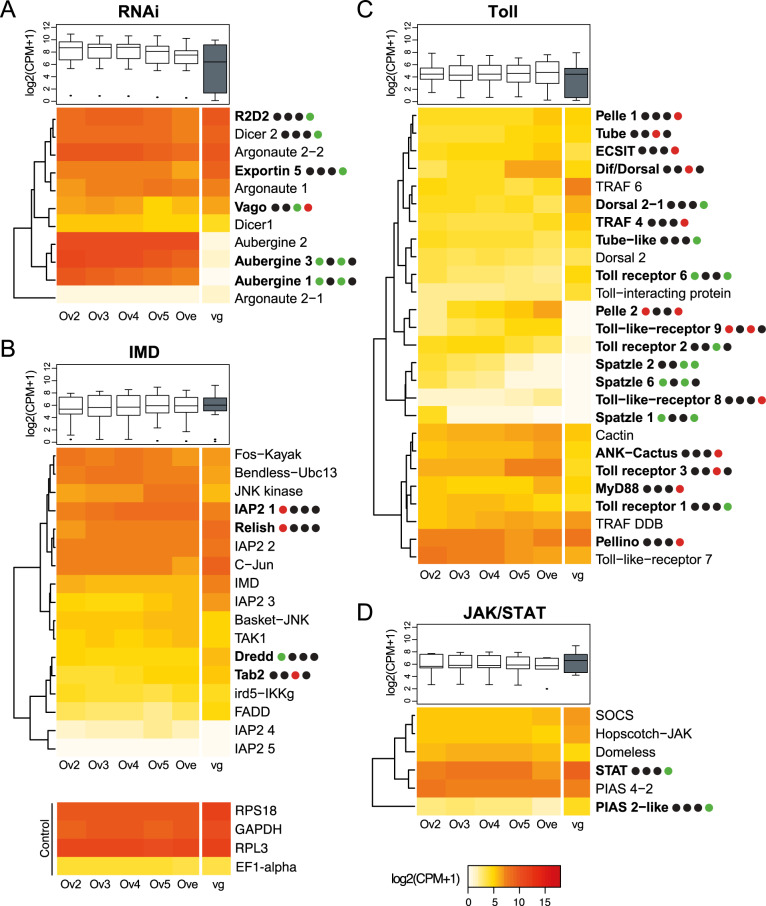
Endogenous viruses form an important proportion of eukaryote genomes and a source of novel functions. How large DNA viruses integrated into a genome evolve when they confer a benefit to their host, however, remains unknown. Bracoviruses are essential for the parasitism success of parasitoid wasps, into whose genomes they integrated ~103 million years ago. Here we show, from the assembly of a parasitoid wasp genome at a chromosomal scale, that bracovirus genes colonized all ten chromosomes of Cotesia congregata. Most form clusters of genes involved in particle production or parasitism success. Genomic comparison with another wasp, Microplitis demolitor, revealed that these clusters were already established ~53 mya and thus belong to remarkably stable genomic structures, the architectures of which are evolutionary constrained. Transcriptomic analyses highlight temporal synchronization of viral gene expression without resulting in immune gene induction, suggesting that no conflicts remain between ancient symbiotic partners when benefits to them converge.
Conflict of interest statement
The authors declare no competing interests.
Figures
References
-
- Parra JRP. biological control in Brazil: an overview. Sci. Agric. 2014;71:420–429.
-
- Dicke M. Behavioural and community ecology of plants that cry for help. Plant Cell Env. 2009;32:654–666. - PubMed
-
- Gauthier J, Drezen JM, Herniou EA. The recurrent domestication of viruses: major evolutionary transitions in parasitic wasps. Parasitology. 2018;145:713–723. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
