

Inducibility, but not stability, of atrial fibrillation is increased by NOX2 overexpression in mice
- PMID: 33483749
- PMCID: PMC8479801
- DOI: 10.1093/cvr/cvab019
Inducibility, but not stability, of atrial fibrillation is increased by NOX2 overexpression in mice
Abstract
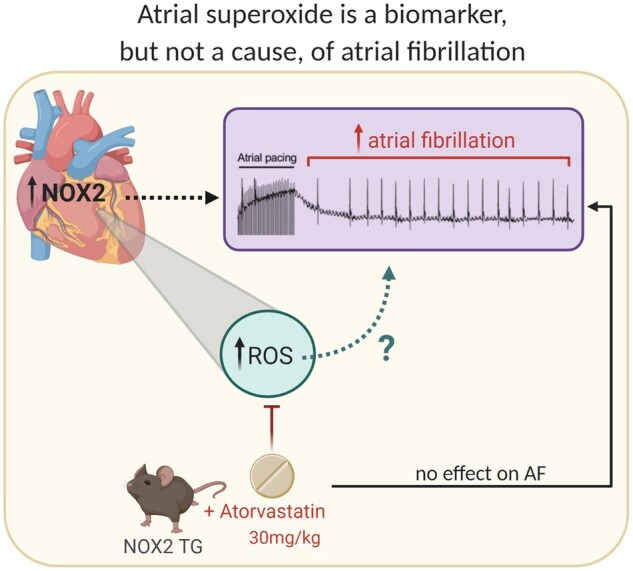
Aims: Gp91-containing NADPH oxidases (NOX2) are a significant source of myocardial superoxide production. An increase in NOX2 activity accompanies atrial fibrillation (AF) induction and electrical remodelling in animal models and predicts incident AF in humans; however, a direct causal role for NOX2 in AF has not been demonstrated. Accordingly, we investigated whether myocardial NOX2 overexpression in mice (NOX2-Tg) is sufficient to generate a favourable substrate for AF and further assessed the effects of atorvastatin, an inhibitor of NOX2, on atrial superoxide production and AF susceptibility.
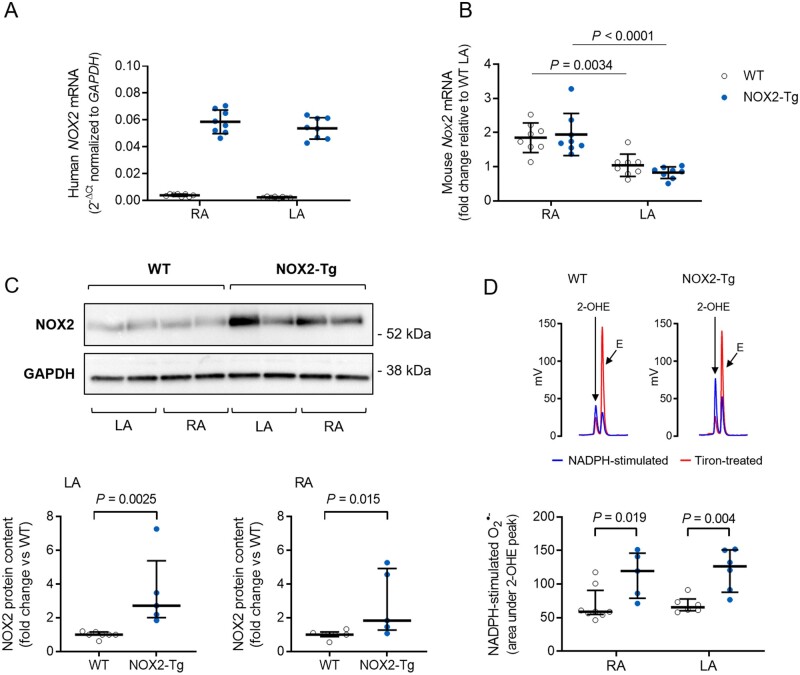
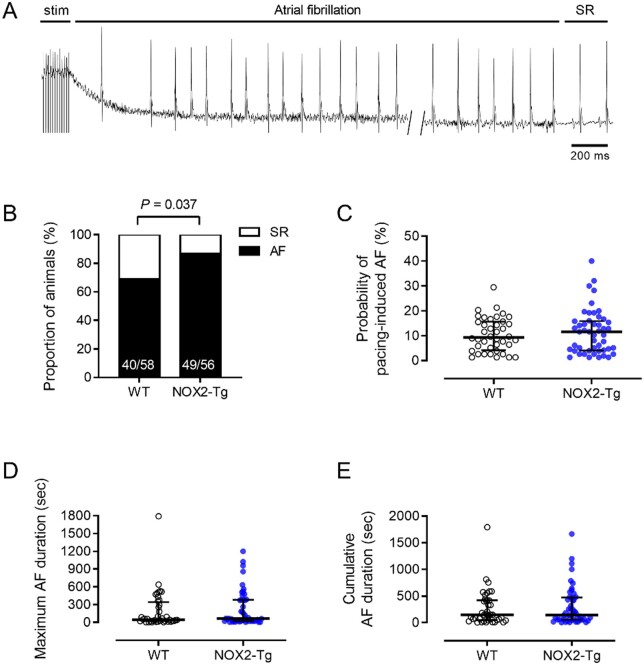
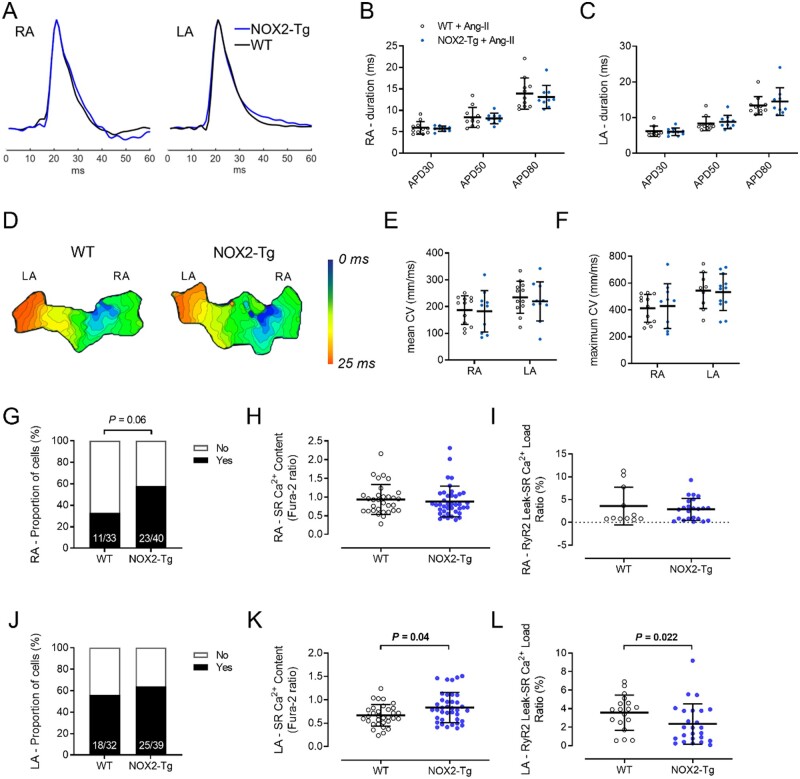
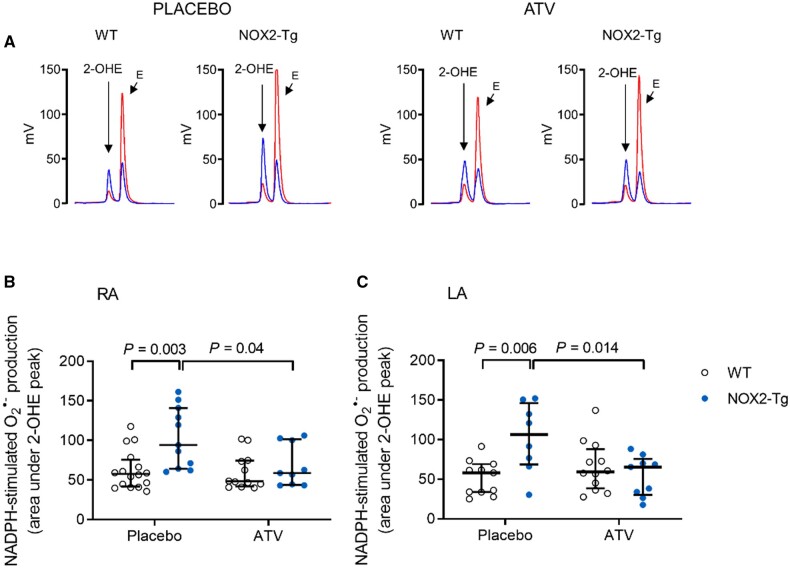
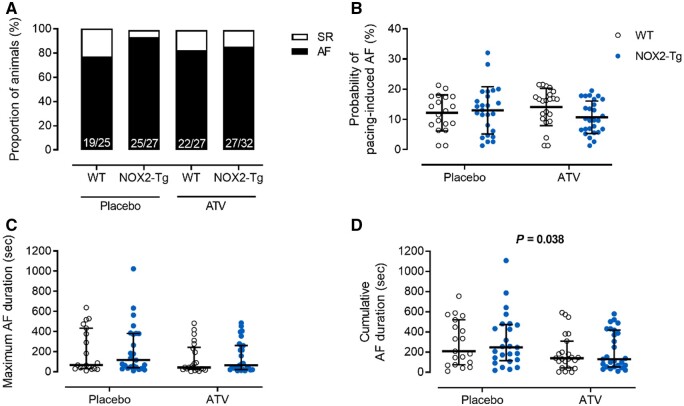
Methods and results: NOX2-Tg mice showed a 2- to 2.5-fold higher atrial protein content of NOX2 compared with wild-type (WT) controls, which was associated with a significant (twofold) increase in NADPH-stimulated superoxide production (2-hydroxyethidium by HPLC) in left and right atrial tissue homogenates (P = 0.004 and P = 0.019, respectively). AF susceptibility assessed in vivo by transoesophageal atrial burst stimulation was modestly increased in NOX2-Tg compared with WT (probability of AF induction: 88% vs. 69%, respectively; P = 0.037), in the absence of significant alterations in AF duration, surface ECG parameters, and LV mass or function. Mechanistic studies did not support a role for NOX2 in promoting electrical or structural remodelling, as high-resolution optical mapping of atrial tissues showed no differences in action potential duration and conduction velocity between genotypes. In addition, we did not observe any genotype difference in markers of fibrosis and inflammation, including atrial collagen content and Col1a1, Il-1β, Il-6, and Mcp-1 mRNA. Similarly, NOX2 overexpression did not have consistent effects on RyR2 Ca2+ leak nor did it affect PKA or CaMKII-mediated RyR2 phosphorylation. Finally, treatment with atorvastatin significantly inhibited atrial superoxide production in NOX2-Tg but had no effect on AF induction in either genotype.
Conclusion: Together, these data indicate that while atrial NOX2 overexpression may contribute to atrial arrhythmogenesis, NOX2-derived superoxide production does not affect the electrical and structural properties of the atrial myocardium.
Keywords: Arrhythmia (mechanisms); Atorvastatin; Atrial fibrillation; NADPH oxidases; Oxidant stress.
© The Author(s) 2021. Published by Oxford University Press on behalf of the European Society of Cardiology.
Figures
Comment in
-
Oxidative stress: a bystander or a causal contributor to atrial remodelling and fibrillation?Cardiovasc Res. 2021 Sep 28;117(11):2291-2293. doi: 10.1093/cvr/cvab124. Cardiovasc Res. 2021. PMID: 33822005 Free PMC article. No abstract available.
References
-
- Chugh SS, Havmoeller R, Narayanan K, Singh D, Rienstra M, Benjamin EJ, Gillum RF, Kim YH, McAnulty JH Jr, Zheng ZJ, Forouzanfar MH, Naghavi M, Mensah GA, Ezzati M, Murray CJ.. Worldwide epidemiology of atrial fibrillation: a Global Burden of Disease 2010 Study. Circulation 2014;129:837–847. - PMC - PubMed
-
- Delgado V, Di Biase L, Leung M, Romero J, Tops LF, Casadei B, Marrouche N, Bax JJ.. Structure and function of the left atrium and left atrial appendage: AF and stroke implications. J Am Coll Cardiol 2017;70:3157–3172. - PubMed
-
- Carnes CA, Chung MK, Nakayama T, Nakayama H, Baliga RS, Piao S, Kanderian A, Pavia S, Hamlin RL, McCarthy PM, Bauer JA, Van Wagoner DR.. Ascorbate attenuates atrial pacing-induced peroxynitrite formation and electrical remodeling and decreases the incidence of postoperative atrial fibrillation. Circ Res 2001;89:e32–e38. - PubMed
-
- Mihm MJ, Yu F, Carnes CA, Reiser PJ, McCarthy PM, Van Wagoner DR, Bauer JA.. Impaired myofibrillar energetics and oxidative injury during human atrial fibrillation. Circulation 2001;104:174–180. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Miscellaneous
