

RNA architecture influences plant biology
- PMID: 33484251
- PMCID: PMC8130982
- DOI: 10.1093/jxb/erab030
RNA architecture influences plant biology
Abstract
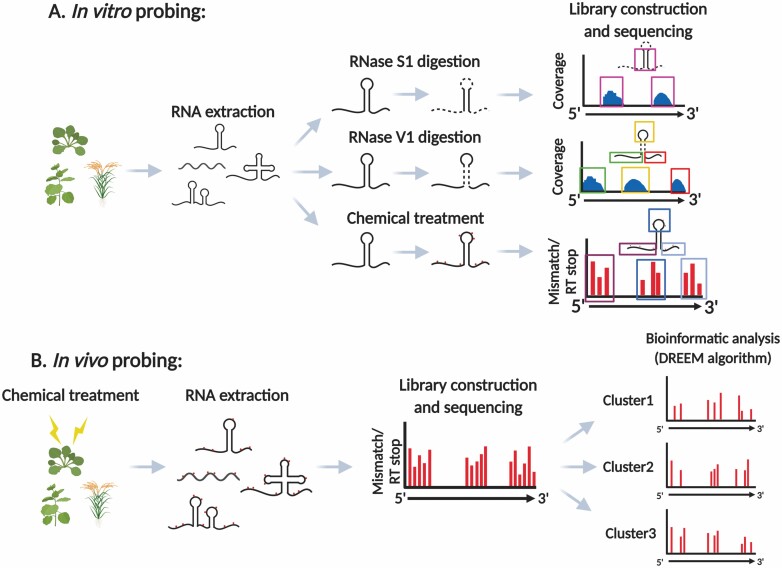
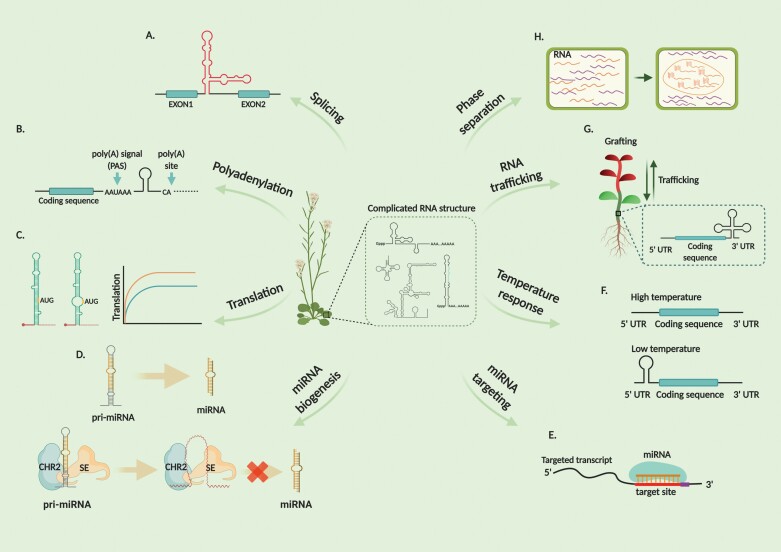
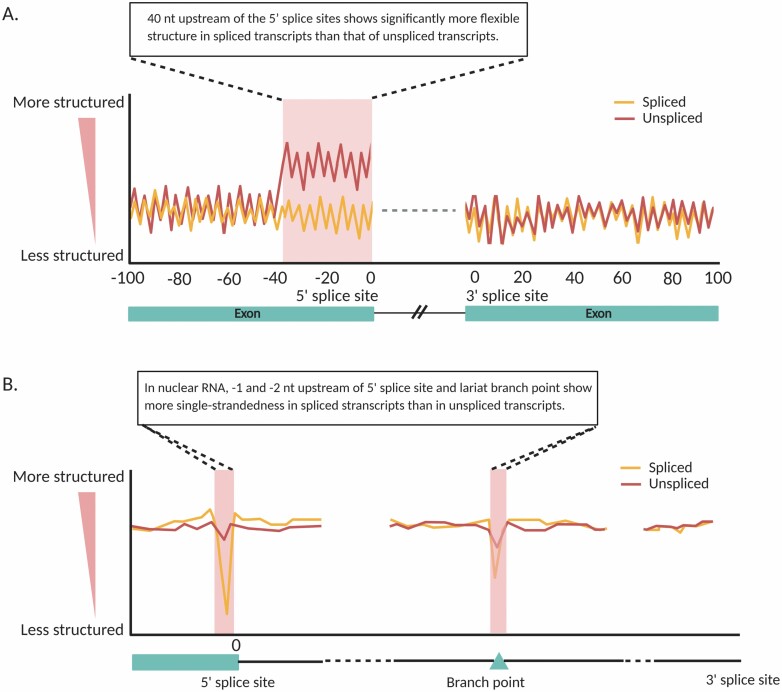
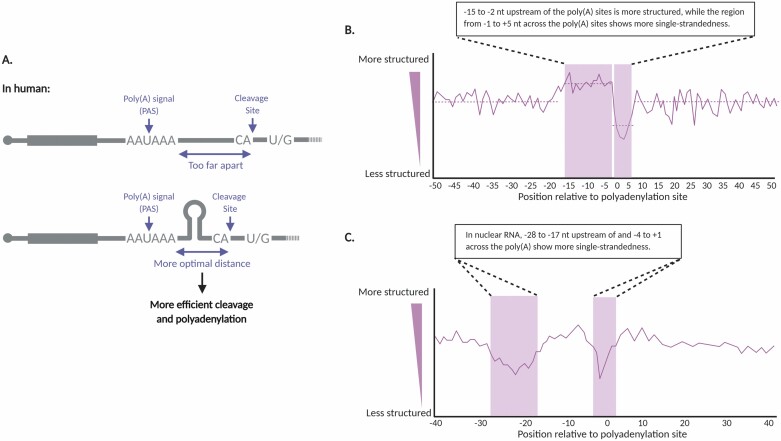
The majority of the genome is transcribed to RNA in living organisms. RNA transcripts can form astonishing arrays of secondary and tertiary structures via Watson-Crick, Hoogsteen, or wobble base pairing. In vivo, RNA folding is not a simple thermodynamic event of minimizing free energy. Instead, the process is constrained by transcription, RNA-binding proteins, steric factors, and the microenvironment. RNA secondary structure (RSS) plays myriad roles in numerous biological processes, such as RNA processing, stability, transportation, and translation in prokaryotes and eukaryotes. Emerging evidence has also implicated RSS in RNA trafficking, liquid-liquid phase separation, and plant responses to environmental variations such as temperature and salinity. At molecular level, RSS is correlated with splicing, polyadenylation, protein synthesis, and miRNA biogenesis and functions. In this review, we summarize newly reported methods for probing RSS in vivo and functions and mechanisms of RSS in plant physiology.
Keywords: RNA secondary structure; RNA trafficking; miRNA; phase separation; polyadenylation; salinity stress; splicing; translation.
© The Author(s) 2021. Published by Oxford University Press on behalf of the Society for Experimental Biology. All rights reserved. For permissions, please email: journals.permissions@oup.com.
Figures
Similar articles
-
Genome-Wide Analysis of RNA Secondary Structure.Annu Rev Genet. 2016 Nov 23;50:235-266. doi: 10.1146/annurev-genet-120215-035034. Epub 2016 Sep 14. Annu Rev Genet. 2016. PMID: 27648642 Review.
-
In vivo genome-wide profiling of RNA secondary structure reveals novel regulatory features.Nature. 2014 Jan 30;505(7485):696-700. doi: 10.1038/nature12756. Epub 2013 Nov 24. Nature. 2014. PMID: 24270811
-
Transcription Regulation Through Nascent RNA Folding.J Mol Biol. 2021 Jul 9;433(14):166975. doi: 10.1016/j.jmb.2021.166975. Epub 2021 Apr 1. J Mol Biol. 2021. PMID: 33811916 Free PMC article. Review.
-
Watson-Crick versus Hoogsteen Base Pairs: Chemical Strategy to Encode and Express Genetic Information in Life.Acc Chem Res. 2021 May 4;54(9):2110-2120. doi: 10.1021/acs.accounts.0c00734. Epub 2021 Feb 16. Acc Chem Res. 2021. PMID: 33591181 Review.
-
Co-Transcriptional RNA Processing in Plants: Exploring from the Perspective of Polyadenylation.Int J Mol Sci. 2021 Mar 24;22(7):3300. doi: 10.3390/ijms22073300. Int J Mol Sci. 2021. PMID: 33804866 Free PMC article. Review.
Cited by
-
Beyond transcription: compelling open questions in plant RNA biology.Plant Cell. 2023 May 29;35(6):1626-1653. doi: 10.1093/plcell/koac346. Plant Cell. 2023. PMID: 36477566 Free PMC article.
-
SlRBP1 promotes translational efficiency via SleIF4A2 to maintain chloroplast function in tomato.Plant Cell. 2022 Jul 4;34(7):2747-2764. doi: 10.1093/plcell/koac104. Plant Cell. 2022. PMID: 35385118 Free PMC article.
-
Structured 3' UTRs destabilize mRNAs in plants.Genome Biol. 2024 Feb 22;25(1):54. doi: 10.1186/s13059-024-03186-x. Genome Biol. 2024. PMID: 38388963 Free PMC article.
-
Recent advances in RNA structurome.Sci China Life Sci. 2022 Jul;65(7):1285-1324. doi: 10.1007/s11427-021-2116-2. Epub 2022 Jun 14. Sci China Life Sci. 2022. PMID: 35717434 Free PMC article. Review.
-
Controlled RISC loading efficiency of miR168 defined by miRNA duplex structure adjusts ARGONAUTE1 homeostasis.Nucleic Acids Res. 2021 Dec 16;49(22):12912-12928. doi: 10.1093/nar/gkab1138. Nucleic Acids Res. 2021. PMID: 34850097 Free PMC article.
References
-
- Anderson SJ, Kramer MC, Gosai SJ, et al. . 2018. N6-methyladenosine inhibits local ribonucleolytic cleavage to stabilize mRNAs in Arabidopsis. Cell Reports 25, 1146–1157. - PubMed
-
- Aw JGA, Lim SW, Wang JX, et al. . 2020. Determination of isoform-specific RNA structure with nanopore long reads. Nature Biotechnology. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
