

Binocular integration of retinal motion information underlies optic flow processing by the cortex
- PMID: 33484637
- PMCID: PMC7987724
- DOI: 10.1016/j.cub.2020.12.034
Binocular integration of retinal motion information underlies optic flow processing by the cortex
Abstract
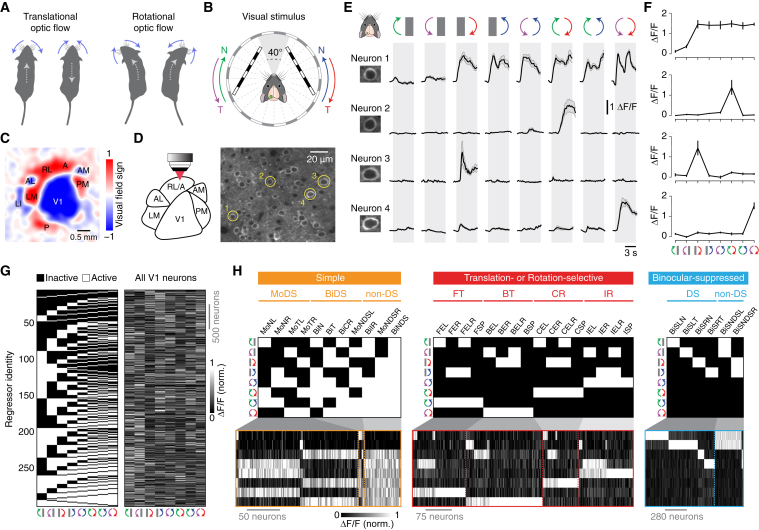
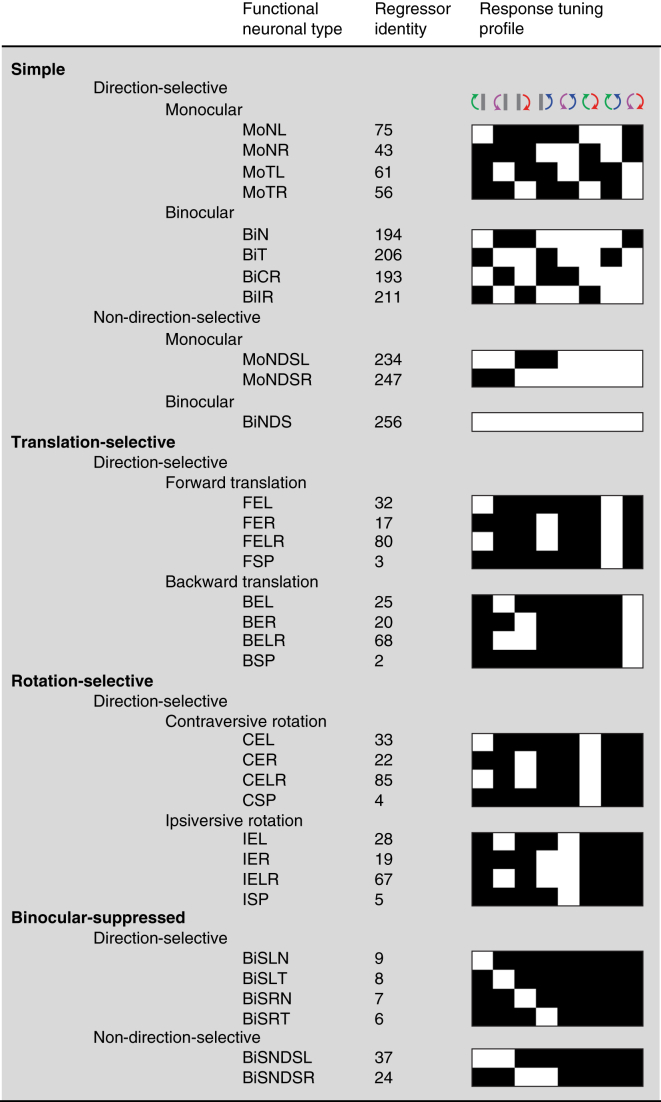
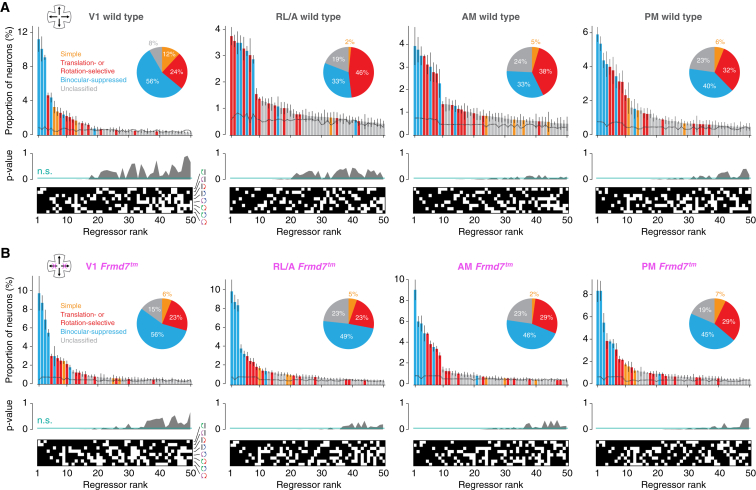
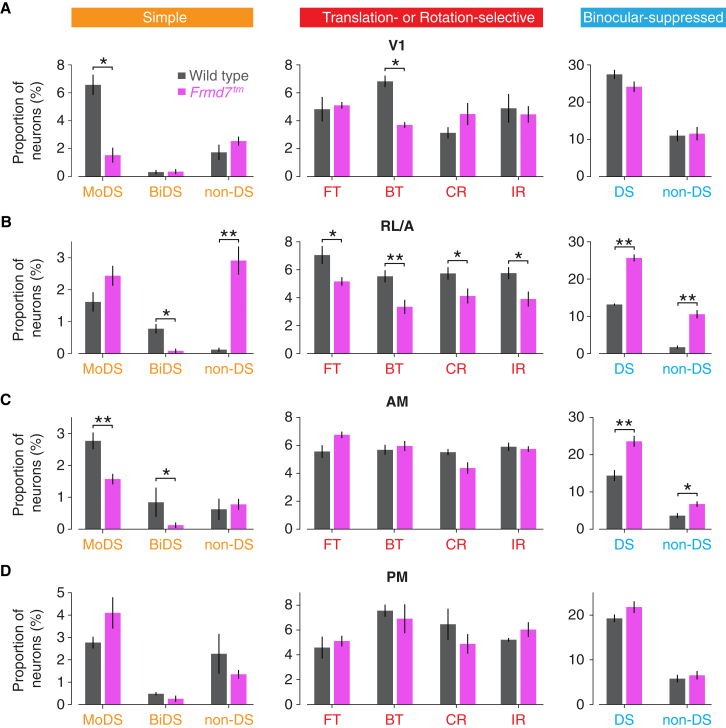
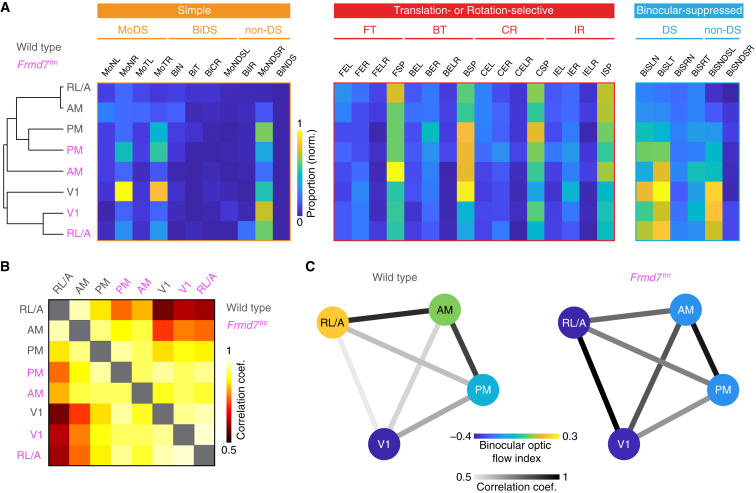
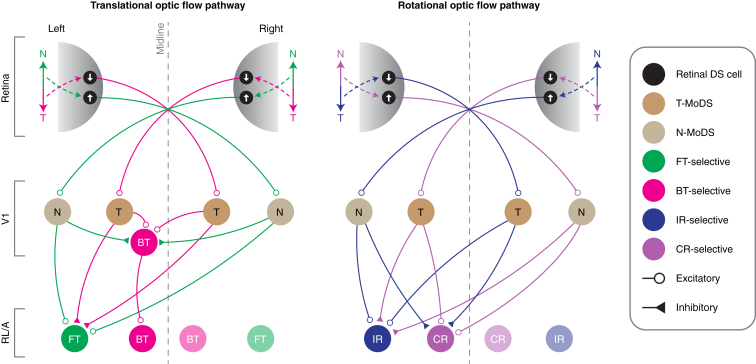
Locomotion creates various patterns of optic flow on the retina, which provide the observer with information about their movement relative to the environment. However, it is unclear how these optic flow patterns are encoded by the cortex. Here, we use two-photon calcium imaging in awake mice to systematically map monocular and binocular responses to horizontal motion in four areas of the visual cortex. We find that neurons selective to translational or rotational optic flow are abundant in higher visual areas, whereas neurons suppressed by binocular motion are more common in the primary visual cortex. Disruption of retinal direction selectivity in Frmd7 mutant mice reduces the number of translation-selective neurons in the primary visual cortex and translation- and rotation-selective neurons as well as binocular direction-selective neurons in the rostrolateral and anterior visual cortex, blurring the functional distinction between primary and higher visual areas. Thus, optic flow representations in specific areas of the visual cortex rely on binocular integration of motion information from the retina.
Keywords: direction-selective cells; intrinsic signal optical imaging; optic flow; retina; rotation; translation; two-photon calcium imaging; visual cortex.
Copyright © 2020 The Author(s). Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests The authors declare no competing interests.
Figures
References
-
- Krapp H.G., Hengstenberg R., Egelhaaf M. Binocular contributions to optic flow processing in the fly visual system. J. Neurophysiol. 2001;85:724–734. - PubMed
-
- Farrow K., Haag J., Borst A. Nonlinear, binocular interactions underlying flow field selectivity of a motion-sensitive neuron. Nat. Neurosci. 2006;9:1312–1320. - PubMed
-
- Kubo F., Hablitzel B., Dal Maschio M., Driever W., Baier H., Arrenberg A.B. Functional architecture of an optic flow-responsive area that drives horizontal eye movements in zebrafish. Neuron. 2014;81:1344–1359. - PubMed
-
- Wylie D.R.W., Bischof W.F., Frost B.J. Common reference frame for neural coding of translational and rotational optic flow. Nature. 1998;392:278–282. - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
