

Four cellulose-active lytic polysaccharide monooxygenases from Cellulomonas species
- PMID: 33485381
- PMCID: PMC7828015
- DOI: 10.1186/s13068-020-01860-3
Four cellulose-active lytic polysaccharide monooxygenases from Cellulomonas species
Abstract
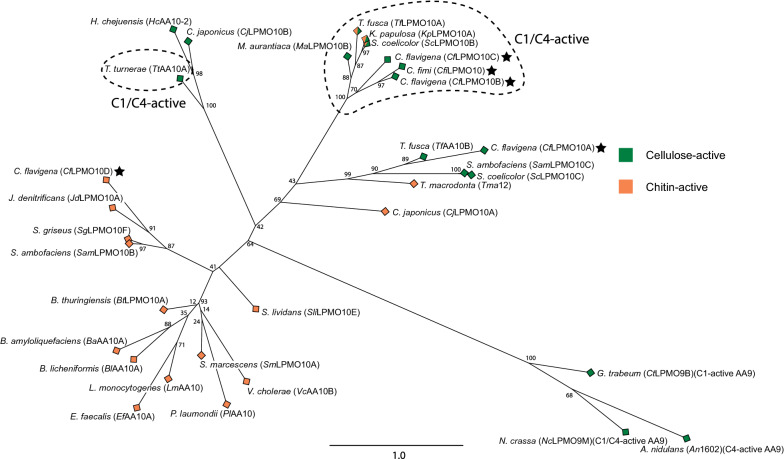
Background: The discovery of lytic polysaccharide monooxygenases (LPMOs) has fundamentally changed our understanding of microbial lignocellulose degradation. Cellulomonas bacteria have a rich history of study due to their ability to degrade recalcitrant cellulose, yet little is known about the predicted LPMOs that they encode from Auxiliary Activity Family 10 (AA10).
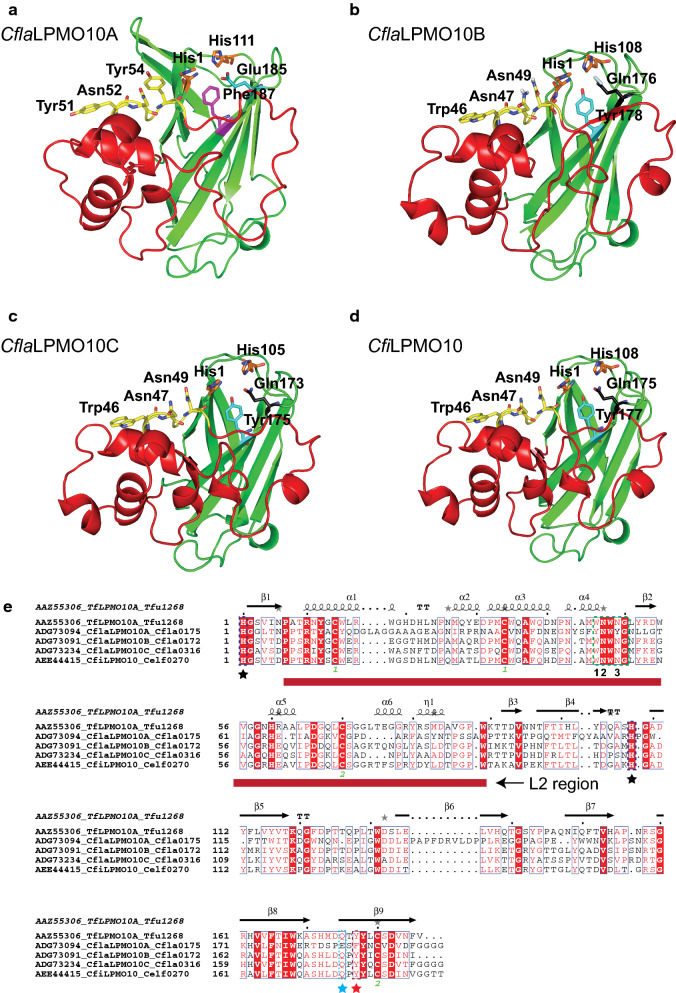
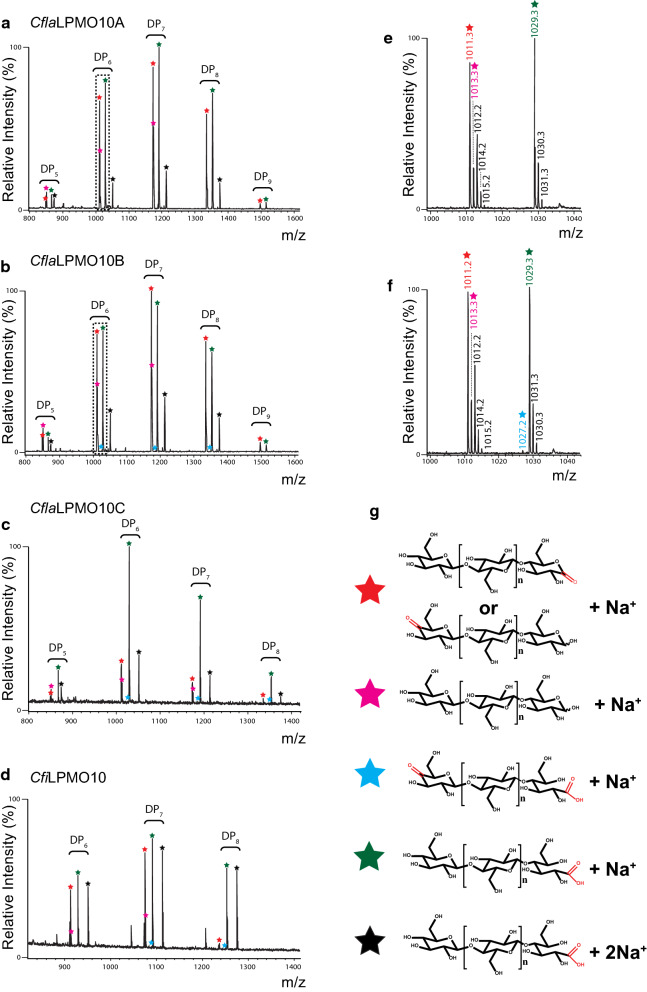
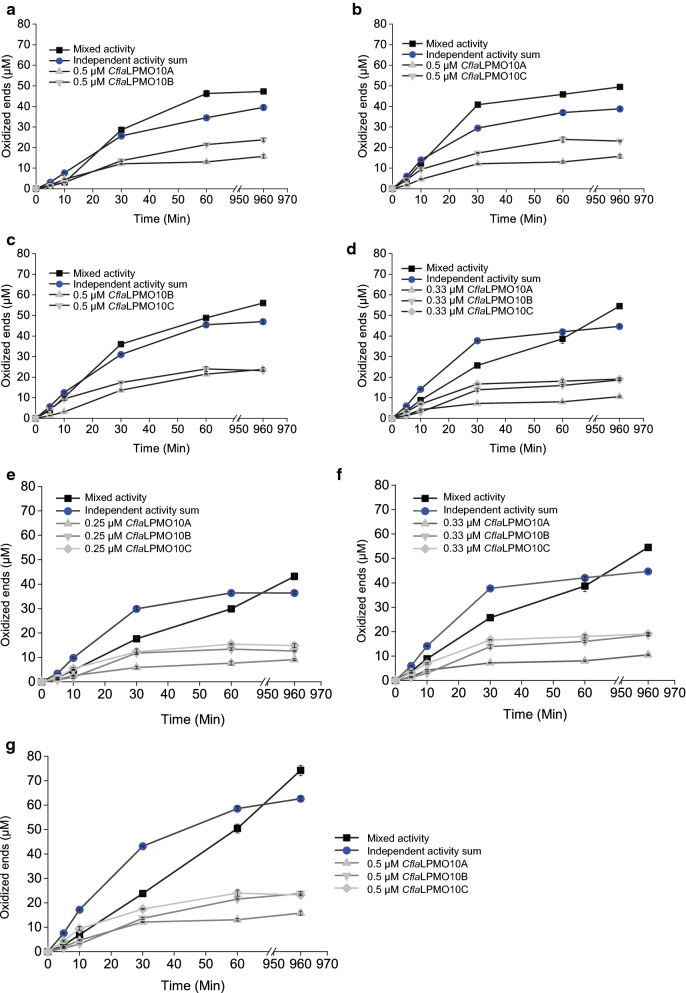
Results: Here, we present the comprehensive biochemical characterization of three AA10 LPMOs from Cellulomonas flavigena (CflaLPMO10A, CflaLPMO10B, and CflaLPMO10C) and one LPMO from Cellulomonas fimi (CfiLPMO10). We demonstrate that these four enzymes oxidize insoluble cellulose with C1 regioselectivity and show a preference for substrates with high surface area. In addition, CflaLPMO10B, CflaLPMO10C, and CfiLPMO10 exhibit limited capacity to perform mixed C1/C4 regioselective oxidative cleavage. Thermostability analysis indicates that these LPMOs can refold spontaneously following denaturation dependent on the presence of copper coordination. Scanning and transmission electron microscopy revealed substrate-specific surface and structural morphological changes following LPMO action on Avicel and phosphoric acid-swollen cellulose (PASC). Further, we demonstrate that the LPMOs encoded by Cellulomonas flavigena exhibit synergy in cellulose degradation, which is due in part to decreased autoinactivation.
Conclusions: Together, these results advance understanding of the cellulose utilization machinery of historically important Cellulomonas species beyond hydrolytic enzymes to include lytic cleavage. This work also contributes to the broader mapping of enzyme activity in Auxiliary Activity Family 10 and provides new biocatalysts for potential applications in biomass modification.
Keywords: AA10; Bioethanol; Biomass; Bioproducts; Cellulose; LPMO.
Conflict of interest statement
The authors declare that they have no competing interests.
Figures



References
-
- Bidlack J, Malone M, Benson R. Molecular structure and component integration of secondary cell walls in plants. Proc Oklahoma Acad Sci. 1992;72:51–56.
-
- Obeng EM, Adam SNN, Budiman C, Ongkudon CM, Maas R, Jose J. Lignocellulases: a review of emerging and developing enzymes, systems, and practices. Bioresour Bioprocessing. 2017 doi: 10.1186/s40643-017-0146-8. - DOI
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
