

Origins of eukaryotic excitability
- PMID: 33487111
- PMCID: PMC7935092
- DOI: 10.1098/rstb.2019.0758
Origins of eukaryotic excitability
Abstract
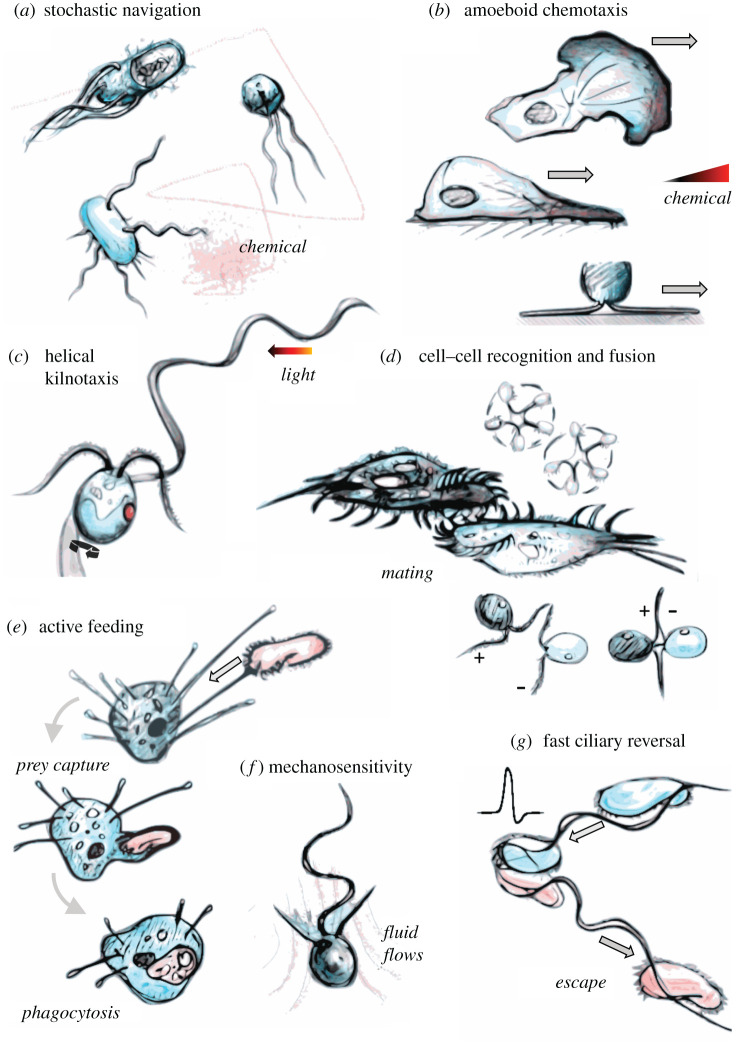
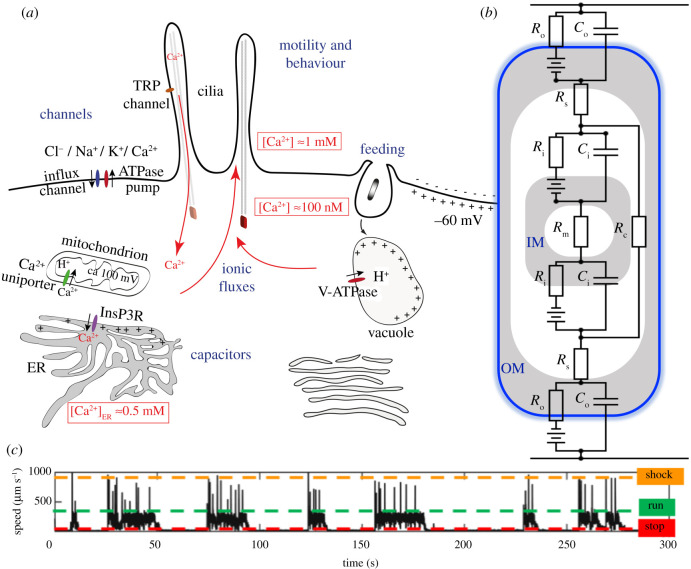
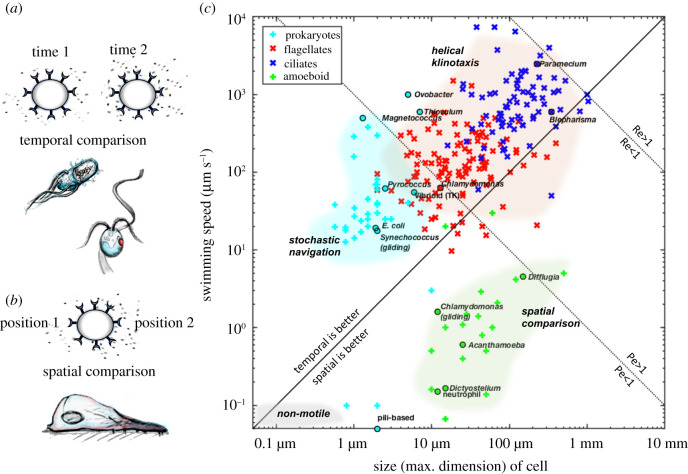
All living cells interact dynamically with a constantly changing world. Eukaryotes, in particular, evolved radically new ways to sense and react to their environment. These advances enabled new and more complex forms of cellular behaviour in eukaryotes, including directional movement, active feeding, mating, and responses to predation. But what are the key events and innovations during eukaryogenesis that made all of this possible? Here we describe the ancestral repertoire of eukaryotic excitability and discuss five major cellular innovations that enabled its evolutionary origin. The innovations include a vastly expanded repertoire of ion channels, the emergence of cilia and pseudopodia, endomembranes as intracellular capacitors, a flexible plasma membrane and the relocation of chemiosmotic ATP synthesis to mitochondria, which liberated the plasma membrane for more complex electrical signalling involved in sensing and reacting. We conjecture that together with an increase in cell size, these new forms of excitability greatly amplified the degrees of freedom associated with cellular responses, allowing eukaryotes to vastly outperform prokaryotes in terms of both speed and accuracy. This comprehensive new perspective on the evolution of excitability enriches our view of eukaryogenesis and emphasizes behaviour and sensing as major contributors to the success of eukaryotes. This article is part of the theme issue 'Basal cognition: conceptual tools and the view from the single cell'.
Keywords: cilia; eukaryogenesis; excitability; membranes; motility; protists.
Figures
References
-
- Izhikevich EM. 2010. Dynamical systems in neuroscience: the geometry of excitability and bursting. Cambridge, MA: MIT Press.
-
- Poincaré H. 1902. La science et l'hypothese [Science and hypothesis], ch. IX. [In French.] Paris, France: Flammarion.
-
- Dusenbery DB. 1992. Sensory ecology: how organisms acquire and respond to information. New York, NY: W. H. Freeman & Company.
Publication types
MeSH terms
Associated data
LinkOut - more resources
Full Text Sources
Other Literature Sources