

Milestones in Bacillus subtilis sporulation research
- PMID: 33490228
- PMCID: PMC7780723
- DOI: 10.15698/mic2021.01.739
Milestones in Bacillus subtilis sporulation research
Abstract
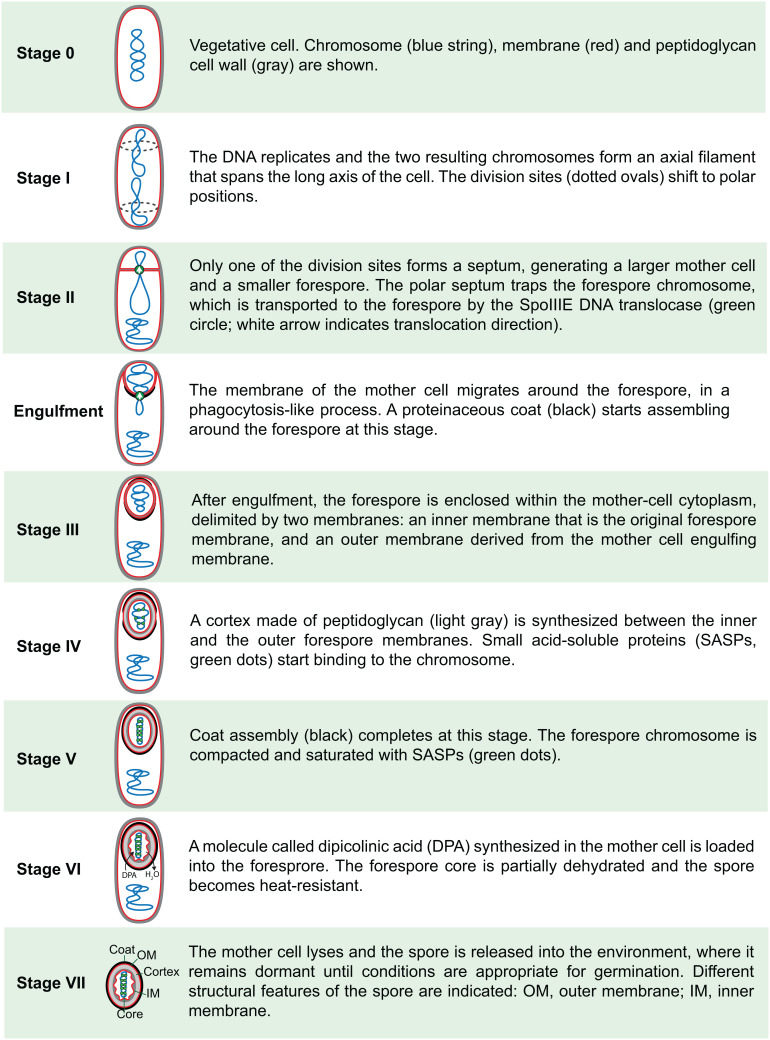
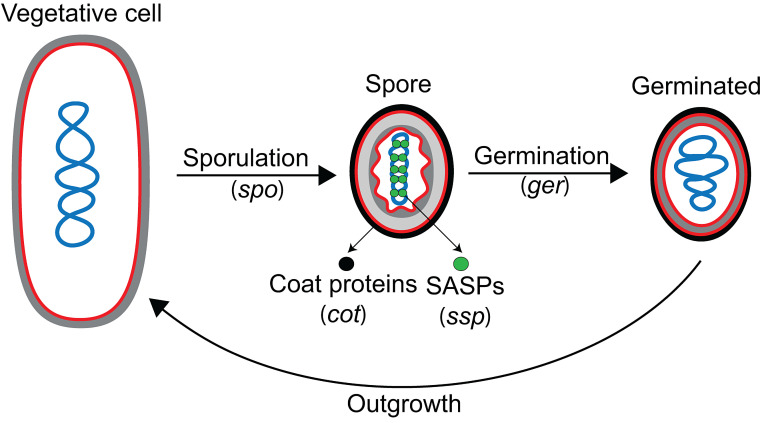
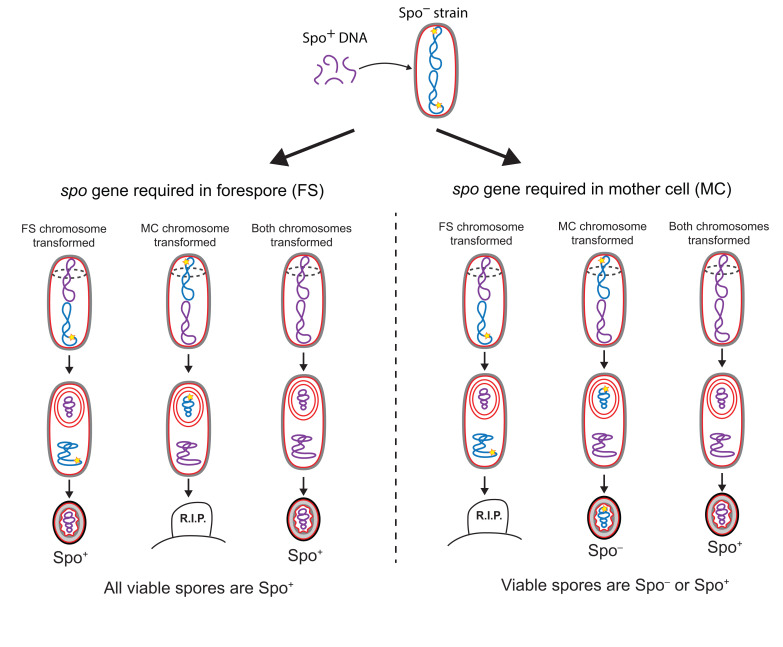
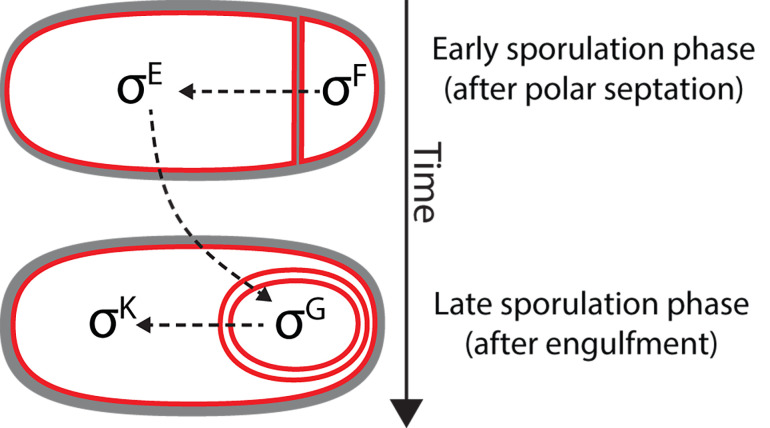
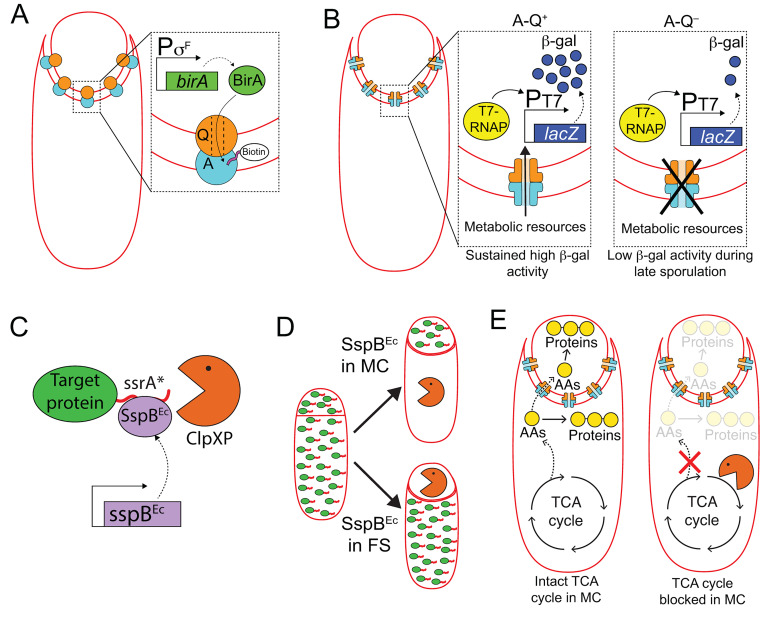
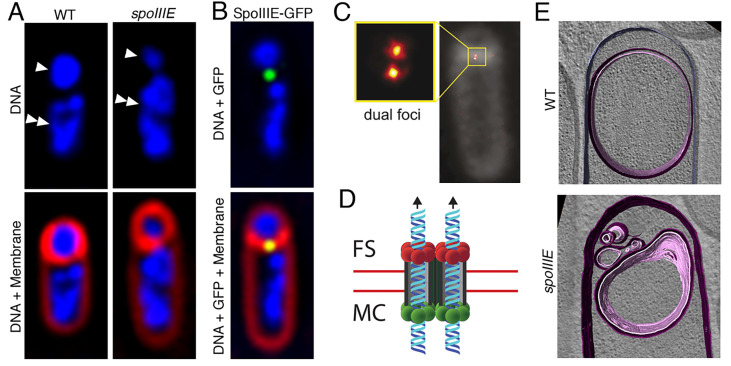
Endospore formation has been a rich field of research for more than a century, and has benefited from the powerful genetic tools available in Bacillus subtilis. In this review, we highlight foundational discoveries that shaped the sporulation field, from its origins to the present day, tracing a chronology that spans more than one hundred eighty years. We detail how cell-specific gene expression has been harnessed to investigate the existence and function of intercellular proteinaceous channels in sporulating cells, and we illustrate the rapid progress in our understanding of the cell biology of sporulation in recent years using the process of chromosome translocation as a storyline. Finally, we sketch general aspects of sporulation that remain largely unexplored, and that we envision will be fruitful areas of future research.
Keywords: Bacillus subtilis; SpoIIIA; SpoIIIE; SpoIIQ; genetics; sporulation; sporulation history.
Copyright: © 2021 Riley et al.
Conflict of interest statement
Conflict of interest: The authors declare no conflict of interest.
Figures
References
-
- Crichton M. Jurassic Park, 1st ed. Alfred A. Knopf. 1990.
-
- Ehrenberg CG. Die Infusionsthierchen als vollkommene Organismen. Verlag Von Leopold Voss, Leipzig. 1838.
-
- Cohn F. Untersuchungen über Bakterien IV. Beiträge zur Biologie der Bazillen. Beitrage zur Biol der Pflanz. 1876. pp. 249–276.
Publication types
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases