

Insights into the Roles of the Sideroflexins/SLC56 Family in Iron Homeostasis and Iron-Sulfur Biogenesis
- PMID: 33494450
- PMCID: PMC7911444
- DOI: 10.3390/biomedicines9020103
Insights into the Roles of the Sideroflexins/SLC56 Family in Iron Homeostasis and Iron-Sulfur Biogenesis
Abstract
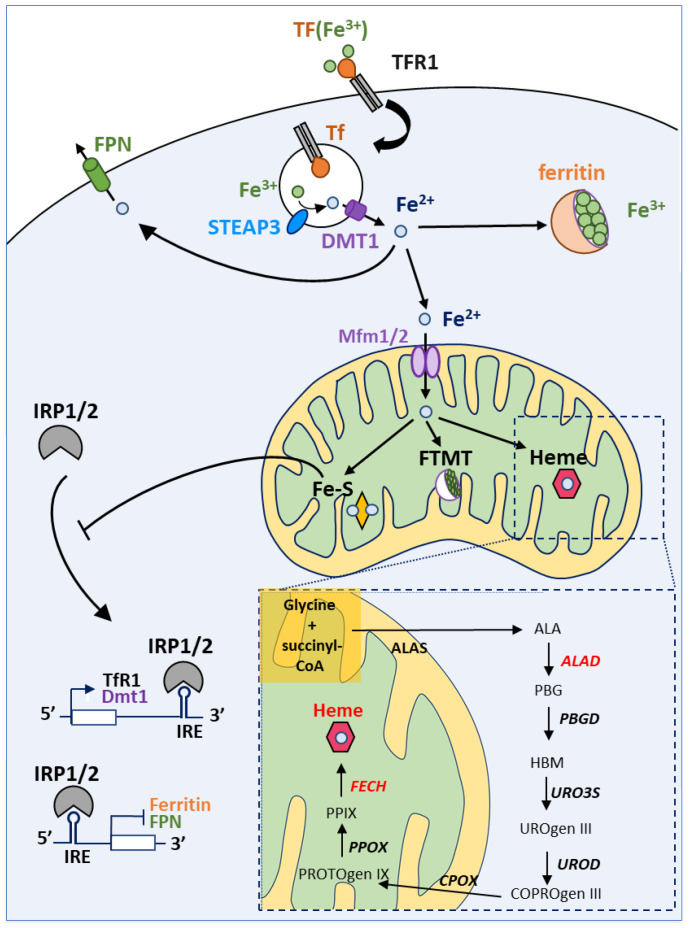
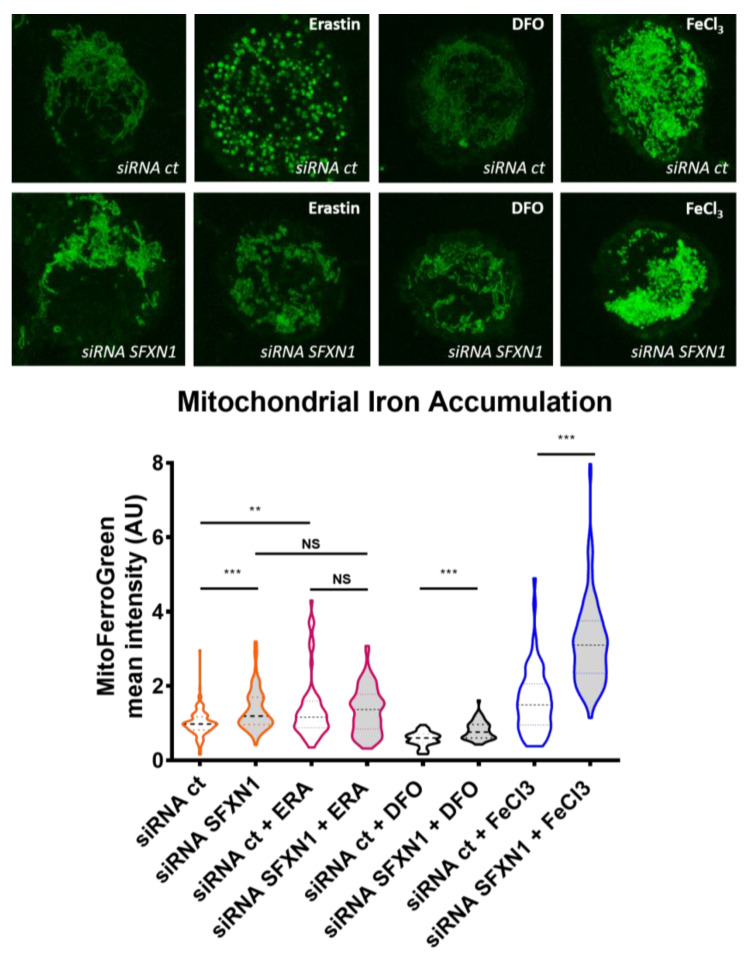
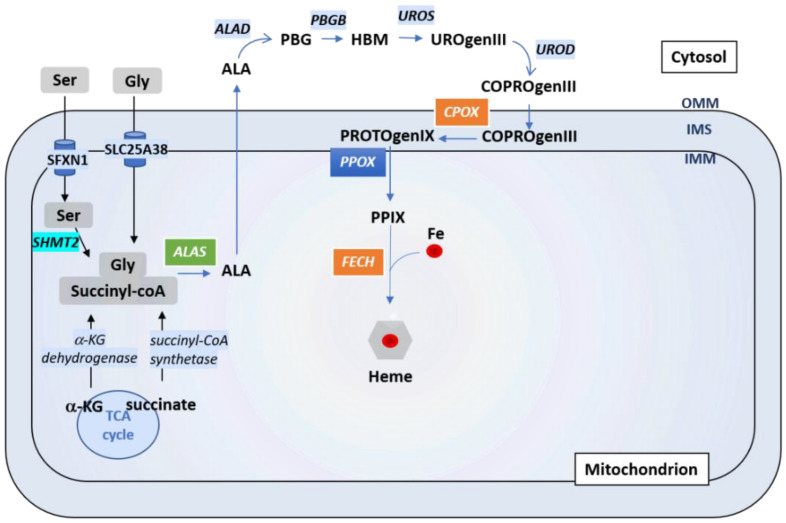
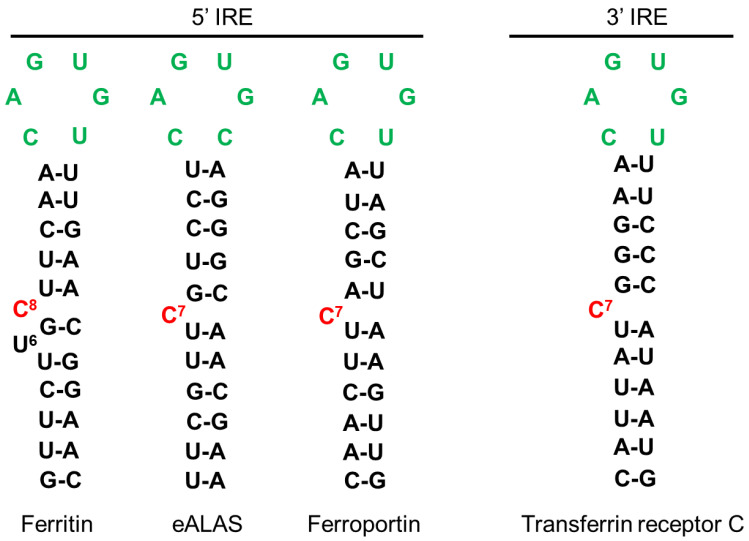
Sideroflexins (SLC56 family) are highly conserved multi-spanning transmembrane proteins inserted in the inner mitochondrial membrane in eukaryotes. Few data are available on their molecular function, but since their first description, they were thought to be metabolite transporters probably required for iron utilization inside the mitochondrion. Such as numerous mitochondrial transporters, sideroflexins remain poorly characterized. The prototypic member SFXN1 has been recently identified as the previously unknown mitochondrial transporter of serine. Nevertheless, pending questions on the molecular function of sideroflexins remain unsolved, especially their link with iron metabolism. Here, we review the current knowledge on sideroflexins, their presumed mitochondrial functions and the sparse-but growing-evidence linking sideroflexins to iron homeostasis and iron-sulfur cluster biogenesis. Since an imbalance in iron homeostasis can be detrimental at the cellular and organismal levels, we also investigate the relationship between sideroflexins, iron and physiological disorders. Investigating Sideroflexins' functions constitutes an emerging research field of great interest and will certainly lead to the main discoveries of mitochondrial physio-pathology.
Keywords: ferritinophagy; ferroptosis; heme biosynthesis; iron homeostasis; iron-sulfur cluster; mitochondria; mitochondrial transporters; one-carbon metabolism; sideroflexin.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
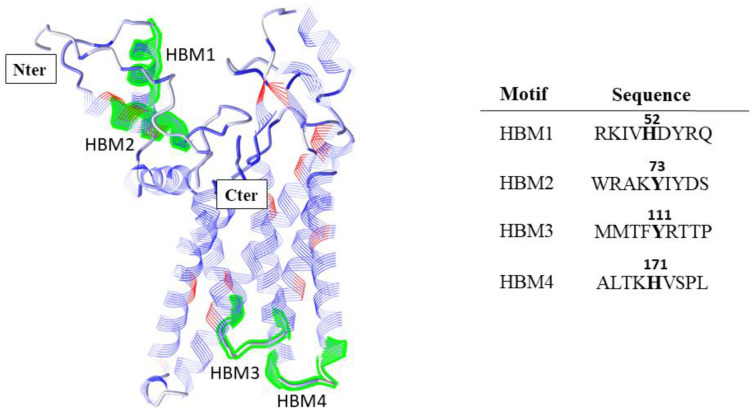
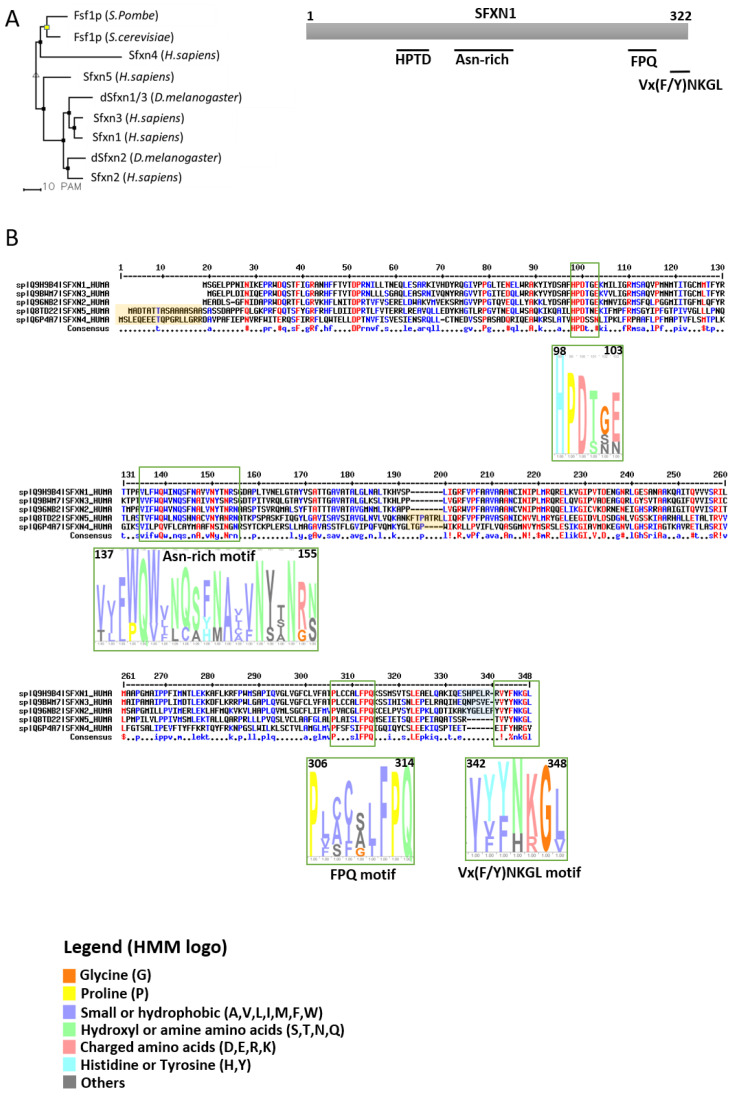
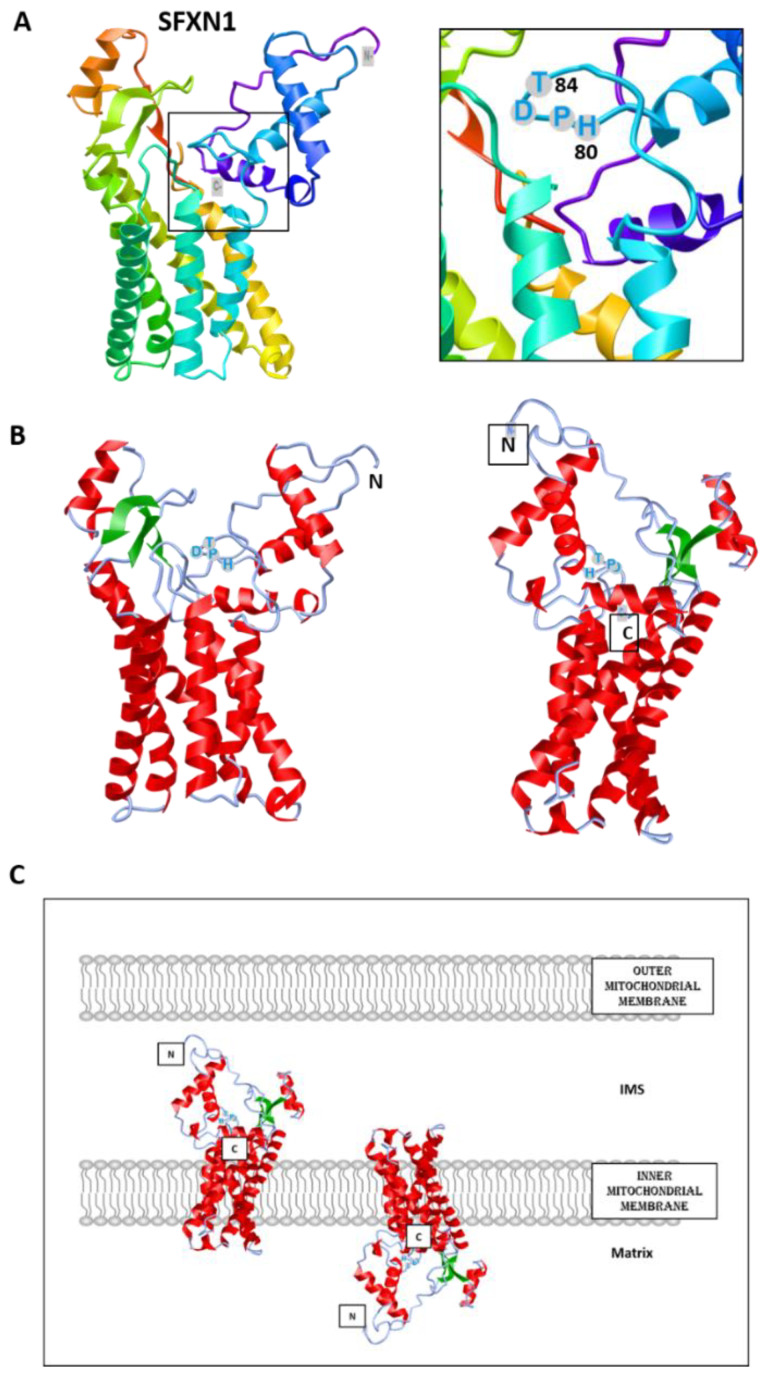
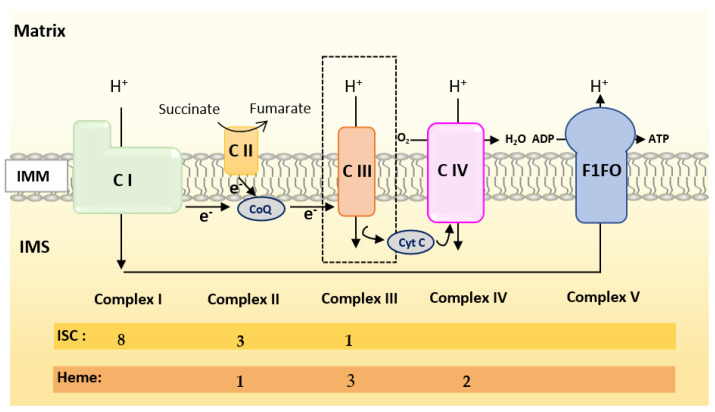
References
-
- SLC56 Sideroflexins IUPHAR/BPS Guide to PHARMACOLOGY. [(accessed on 20 January 2021)];2020 Available online: http://www.guidetopharmacology.org/GRAC/FamilyDisplayForward?familyId=1008.
Publication types
LinkOut - more resources
Full Text Sources
Other Literature Sources
