

Emerging functions of branched ubiquitin chains
- PMID: 33495455
- PMCID: PMC7835216
- DOI: 10.1038/s41421-020-00237-y
Emerging functions of branched ubiquitin chains
Abstract
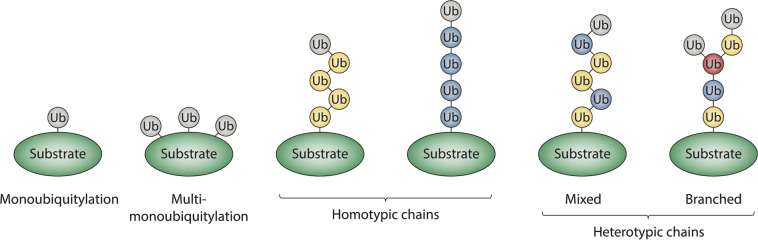
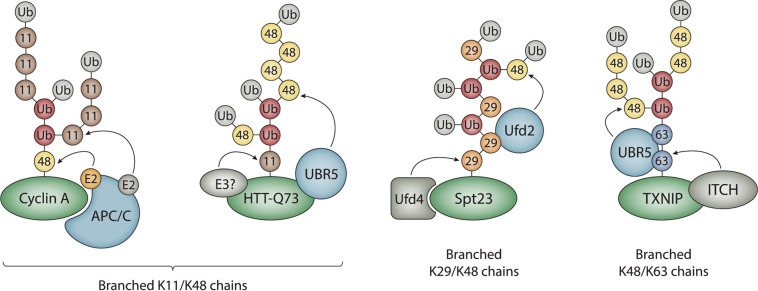
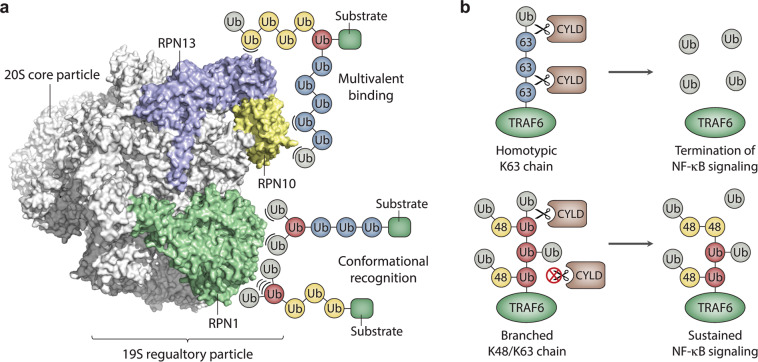
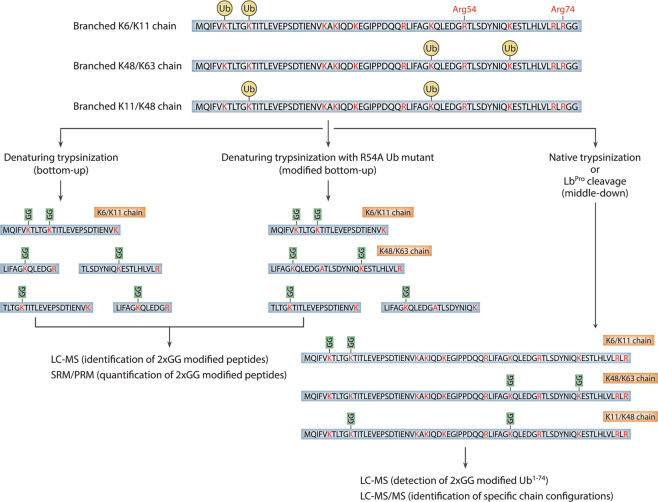
Ubiquitylation is a critical post-translational modification that controls a wide variety of processes in eukaryotes. Ubiquitin chains of different topologies are specialized for different cellular functions and control the stability, activity, interaction properties, and localization of many different proteins. Recent work has highlighted a role for branched ubiquitin chains in the regulation of cell signaling and protein degradation pathways. Similar to their unbranched counterparts, branched ubiquitin chains are remarkably diverse in terms of their chemical linkages, structures, and the biological information they transmit. In this review, we discuss emerging themes related to the architecture, synthesis, and functions of branched ubiquitin chains. We also describe methodologies that have recently been developed to identify and decode the functions of these branched polymers.
Conflict of interest statement
The authors declare that they have no conflict of interest.
Figures
References
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
