

Viral Hijack of Filamentous Surface Structures in Archaea and Bacteria
- PMID: 33499367
- PMCID: PMC7911016
- DOI: 10.3390/v13020164
Viral Hijack of Filamentous Surface Structures in Archaea and Bacteria
Abstract
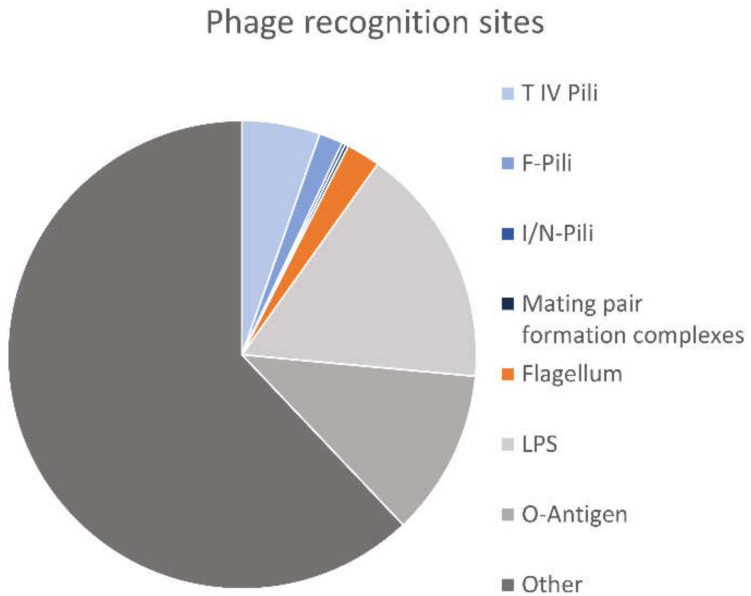
The bacterial and archaeal cell surface is decorated with filamentous surface structures that are used for different functions, such as motility, DNA exchange and biofilm formation. Viruses hijack these structures and use them to ride to the cell surface for successful entry. In this review, we describe currently known mechanisms for viral attachment, translocation, and entry via filamentous surface structures. We describe the different mechanisms used to exploit various surface structures bacterial and archaeal viruses. This overview highlights the importance of filamentous structures at the cell surface for entry of prokaryotic viruses.
Keywords: archaeal virus; archaellum; flagellum; phage; pilus; viral entry.
Conflict of interest statement
The authors declare no conflict of interest.
Figures


References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
