

The yellow perch (Perca flavescens) microbiome revealed resistance to colonisation mostly associated with neutralism driven by rare taxa under cadmium disturbance
- PMID: 33499999
- PMCID: PMC7934398
- DOI: 10.1186/s42523-020-00063-3
The yellow perch (Perca flavescens) microbiome revealed resistance to colonisation mostly associated with neutralism driven by rare taxa under cadmium disturbance
Abstract
Background: Disentangling the dynamics of microbial interactions within communities improves our comprehension of metacommunity assembly of microbiota during host development and under perturbations. To assess the impact of stochastic variation of neutral processes on microbiota structure and composition under disturbance, two types of microbial habitats, free-living (water), and host-associated (skin and gut) were experimentally exposed to either a constant or gradual selection regime exerted by two sublethal cadmium chloride dosages (CdCl2). Yellow Perch (Perca flavescens) was used as a piscivorous ecotoxicological model. Using 16S rDNA gene based metataxonomics, quantitative diversity metrics of water, skin and gut microbial communities were characterized along with development and across experimental conditions.
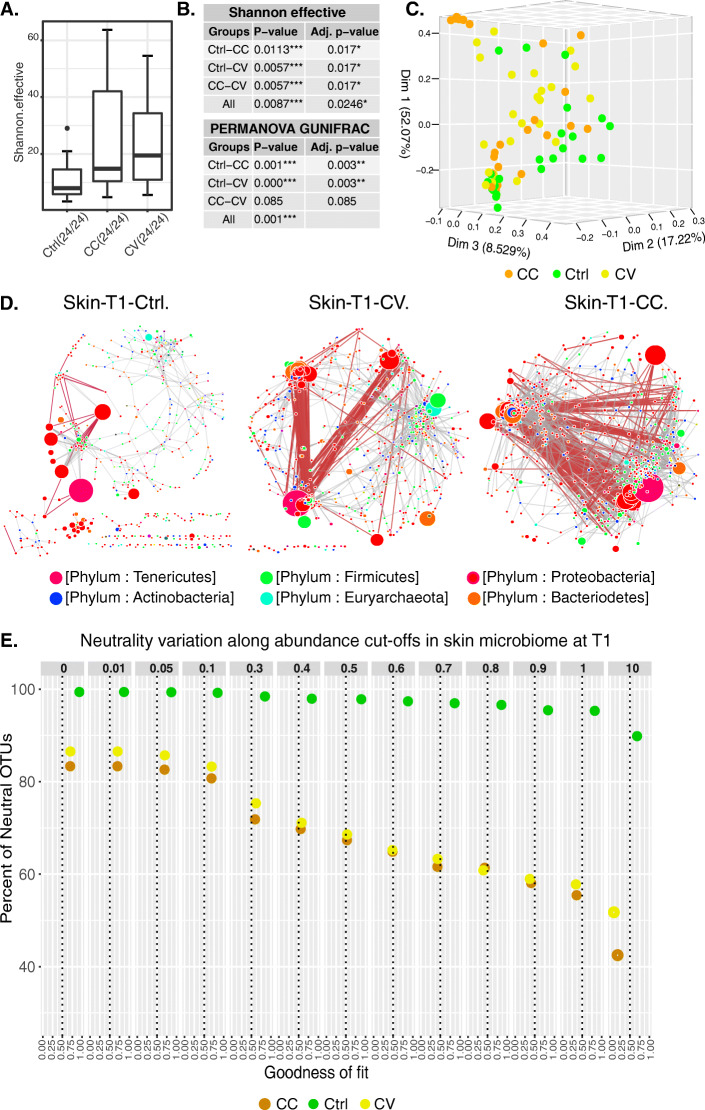
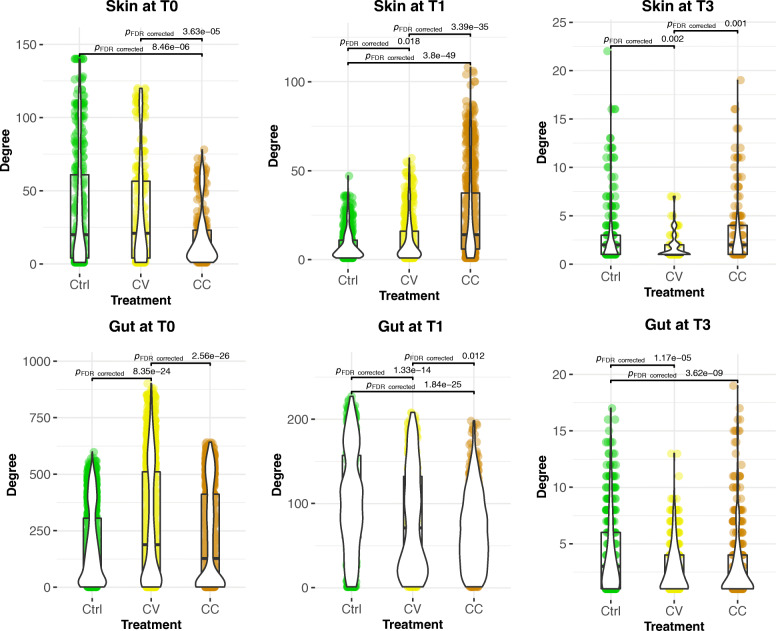
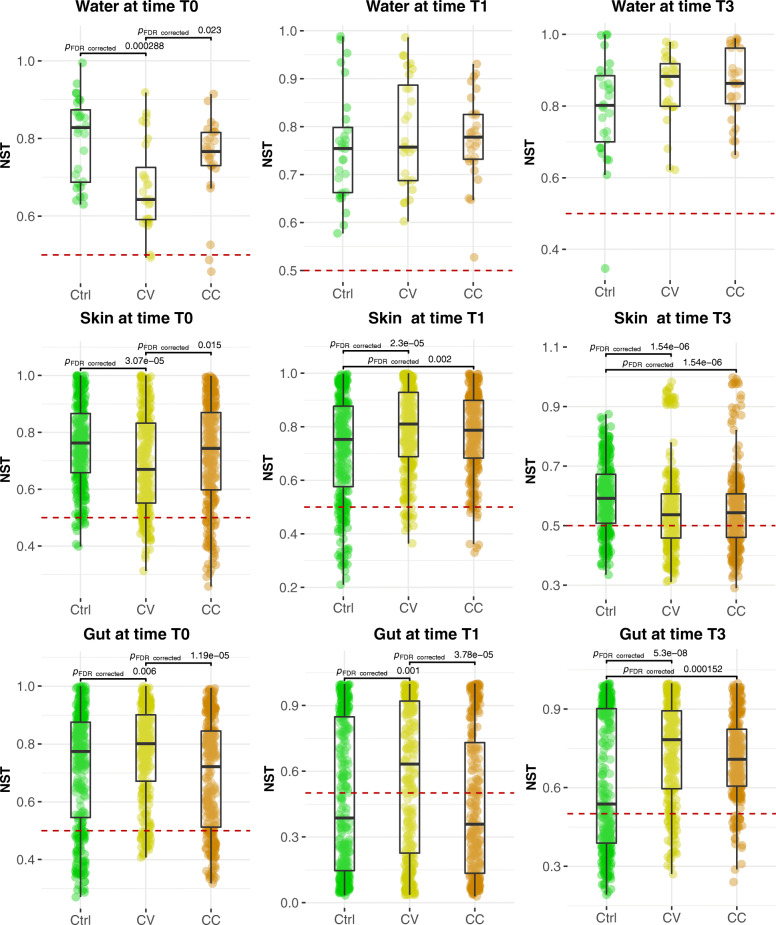
Results: After 30 days, constant and gradual selection regimes drove a significant alpha diversity increase for both skin and gut microbiota. In the skin, pervasive negative correlations between taxa in both selection regimes in addition to the taxonomic convergence with the environmental bacterial community, suggest a loss of colonisation resistance resulting in the dysbiosis of yellow perch microbiota. Furthermore, the network connectivity in gut microbiome was exclusively maintained by rare (low abundance) OTUs, while most abundant OTUs were mainly composed of opportunistic invaders such as Mycoplasma and other genera related to fish pathogens such as Flavobacterium. Finally, the mathematical modelling of community assembly using both non-linear least squares models (NLS) based estimates of migration rates and normalized stochasticity ratios (NST) based beta-diversity distances suggested neutral processes drove by taxonomic drift in host and water communities for almost all treatments. The NLS models predicted higher demographic stochasticity in the cadmium-free host and water microbiomes, however, NST models suggested higher ecological stochasticity under perturbations.
Conclusions: Neutral models agree that water and host-microbiota assembly promoted by rare taxa have evolved predominantly under neutral processes with potential involvement of deterministic forces sourced from host filtering and cadmium selection. The early signals of perturbations in the skin microbiome revealed antagonistic interactions by a preponderance of negative correlations in the co-abundance networks. Our findings enhance our understanding of community assembly host-associated and free-living under anthropogenic selective pressure.
Conflict of interest statement
The authors declare that they have no competing interests.
Figures




References
LinkOut - more resources
Full Text Sources
Other Literature Sources