

Genome diversity and instability in human germ cells and preimplantation embryos
- PMID: 33500205
- PMCID: PMC8097364
- DOI: 10.1016/j.semcdb.2020.12.007
Genome diversity and instability in human germ cells and preimplantation embryos
Abstract
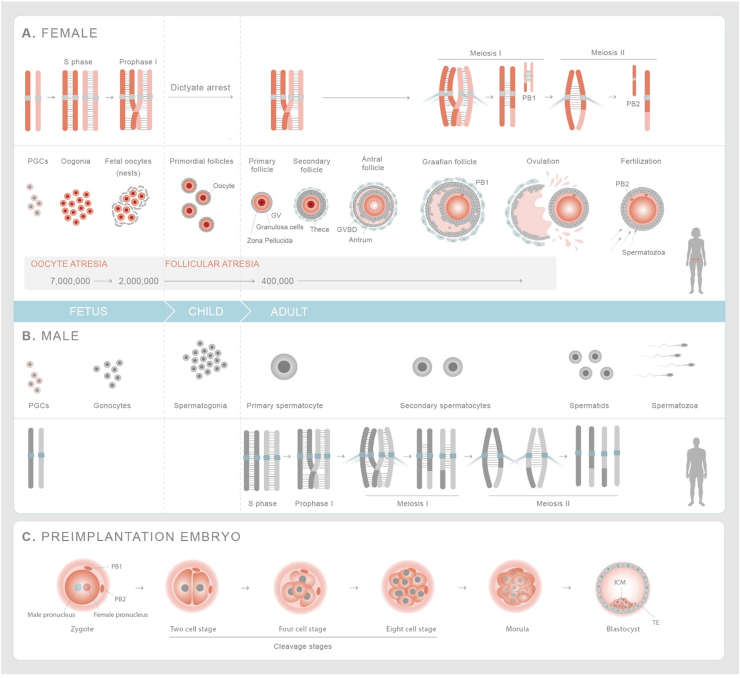
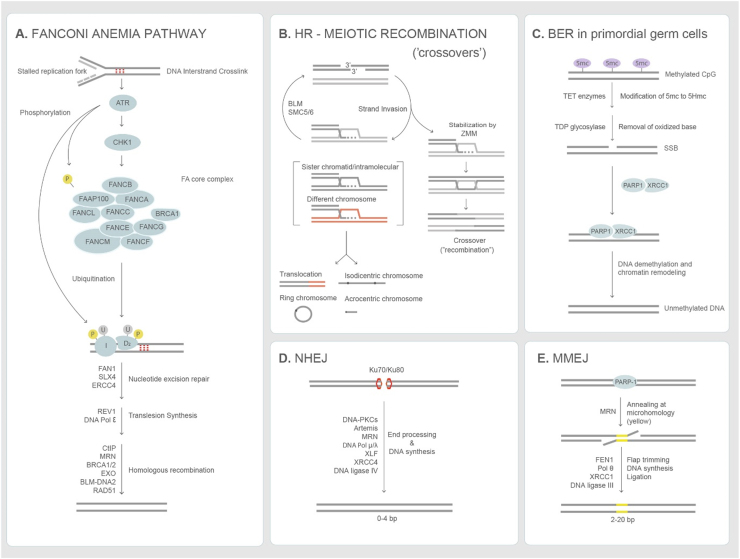
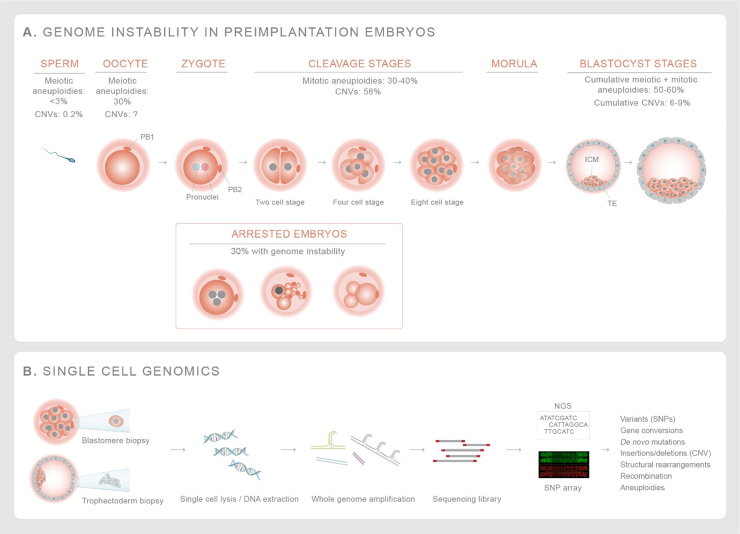
Genome diversity is essential for evolution and is of fundamental importance to human health. Generating genome diversity requires phases of DNA damage and repair that can cause genome instability. Humans have a high incidence of de novo congenital disorders compared to other organisms. Recent access to eggs, sperm and preimplantation embryos is revealing unprecedented rates of genome instability that may result in infertility and de novo mutations that cause genomic imbalance in at least 70% of conceptions. The error type and incidence of de novo mutations differ during developmental stages and are influenced by differences in male and female meiosis. In females, DNA repair is a critical factor that determines fertility and reproductive lifespan. In males, aberrant meiotic recombination causes infertility, embryonic failure and pregnancy loss. Evidence suggest germ cells are remarkably diverse in the type of genome instability that they display and the DNA damage responses they deploy. Additionally, the initial embryonic cell cycles are characterized by a high degree of genome instability that cause congenital disorders and may limit the use of CRISPR-Cas9 for heritable genome editing.
Keywords: Aneuploidy; CNVs; DNA damage response; Genome instability; Genomic disorders; Human oocytes and embryos; Rearrangements.
Copyright © 2021. Published by Elsevier Ltd.
Figures

References
-
- Olivieri M., Cho T., Álvarez-Quilón A., Li K., Schellenberg M.J., Zimmermann M., Hustedt N., Rossi S.E., Adam S., Melo H., Heijink A.M., Sastre-Moreno G., Moatti N., Szilard R.K., McEwan A., Ling A.K., Serrano-Benitez A., Ubhi T., Feng S., Pawling J., Delgado-Sainz I., Ferguson M.W., Dennis J.W., Brown G.W., Cortés-Ledesma F., Williams R.S., Martin A., Xu D., Durocher D. A genetic map of the response to DNA damage in human cells. Cell. 2020;182(2):481–496. e21. - PMC - PubMed
-
- Ford C.E., Jones K.W., Miller O.J., Mittwoch U., Penrose L.S., Ridler M., Shapiro A. The chromosomes in a patient showing both mongolism and the Klinefelter syndrome. Lancet. 1959;1(7075):709–710. - PubMed
-
- Ford C.E., Polani P.E., Briggs J.H., Bishop P.M. A presumptive human XXY/XX mosaic. Nature. 1959;183(4667):1030–1032. - PubMed
-
- Lejeune J., Turpin R., Gautier M. Mongolism; a chromosomal disease (trisomy) Bull. Acad. Natl. Med. 1959;143(11–12):256–265. - PubMed
-
- Jacobs P.A., Baikie A.G., Court Brown W.M., Strong J.A. The somatic chromosomes in mongolism. Lancet. 1959;1(7075):710. - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources
